Pflanzen-assoziierte Mikroben spielen eine wichtige Rolle im biogeochemischen Radfahren, Bioremediation, Klimaschutz, Pflanzenwachstum und Gesundheit sowie Pflanzenverträglichkeit gegenüber biotischen und abiotischen Belastungen. Mikroorganismen interagieren mit Pflanzen sowohl direkt durch den Pflanzenzellwandkontakt als auch indirekt über chemische Sekretion und Signalisierung 1 , 2 , 3 . Als sessile Organismen haben Pflanzen direkte und indirekte Mechanismen entwickelt, um einer Infektion durch Pathogene zu widerstehen. Direkte Abwehr umfasst strukturelle Abwehrkräfte und die Expression von Verteidigungsproteinen, während indirekte Verteidigungen sekundäre Pflanzenmetabolitproduktion und die Anziehung von Organismen antagonistisch gegenüber eindringenden Pathogenen 4 , 5 umfassen . Pflanzen-abgeleitete Wurzel-Exsudate, Sekrete, Schleime, Mucigel und Lysate verändern die physikalisch-chemischen Eigenschaften der Rhizosphäre zu gewinnen oder zu stoßenMikroben gegen ihre Gastgeber 6 . Die chemische Zusammensetzung der Wurzelsekretion ist speziesspezifisch und dient so als selektiver Filter, der es ermöglicht, dass bestimmte Mikroorganismen, die in der Lage sind, solche Verbindungen zu erkennen, in der Rhizosphäre 6 zu blühen. So können kompatible mikrobielle Spezies stimuliert werden, um ihre Assoziationen zu aktivieren und zu verstärken, entweder zum Nutzen oder zum Nachteil des Pflanzenwirts 1 .
Das Verständnis von Pflanzen-Mikroben-Wechselwirkungen in der Rhizosphäre ist der Schlüssel zur Steigerung der Pflanzenproduktivität und des Ökosystems, da eine Mehrheit der mikrobiellen und chemischen Exposition an der Wurzelstruktur und der Boden-Luft-Grenzfläche 2 , 6 , 7 , 8 erfolgt . Allerdings ist die Untersuchung von unterirdischen Pflanzen-Mikroben-Wechselwirkungen und wechselseitigen Reaktionen aufgrund ihrer faszinierenden Herausforderung eine Herausforderung Komplexe und dynamische Natur und der Mangel an geeigneten experimentellen Modellen mit natürlicher Wurzelstruktur und Pflanzenmorphologie unter streng kontrollierbaren Wachstumsbedingungen. Als einer der am stärksten untersuchten Phytopathogene infiziert Agrobacterium eine breite Palette von Pflanzen mit landwirtschaftlicher und gartenbaulicher Bedeutung, darunter Kirsche, Apfel, Birne, Traube und Rose 9 . Agrobacterium ist ein wichtiger Modellorganismus für das Verständnis von Pflanzen-Pathogen-Wechselwirkungen und ist ein leistungsfähiges Werkzeug in der Pflanzen-Transformation und Anlagentechnik 10 , 11 , 12 , 13 , 14 .
Molekulare Pflanzen- Agrobacterium- Wechselwirkungen wurden seit mehreren Jahrzehnten gut untersucht, und das aktuelle Verständnis der Agrobacterium- Pathogenität ist umfangreich 9 ,f "> 11, 15, 16. Agrobacterium Pathogenität in seine entwickelte Fähigkeiten des Wahrnehmens von Pflanzen abgeleiteten Signale weitgehend zurückgeführt, in der feine Modulation seiner Virulenz Programm und Zell-zu-Zell – Kommunikation führt, sogenannte quorum sensing 17. Der Das Agrobacterium- Virulenz-Programm wird durch mehrere in der Rhizosphäre verfügbare Signale reguliert und umfasst zwei Sätze von 2-Komponenten-Systemen, das ChvG / I-System und das VirA / G-System. Säurebedingungen in der Rhizosphäre aktivieren die Transkription von chvG / I , virA / G , Und mehrere andere Gene, die an der Agrobacterium- Pathogenität beteiligt sind, einschließlich virE0 , virE1 , virH1 , virH2 und Genen des Typ-VI-Sekretionssystems (T6SS) 18. Pflanzliche Phenolverbindungen, einschließlich Acetosyringon (4'-hydroxy-3 ', 5 '-Dimethoxyacetophenon), aktivieren Sie die V.IrA / G 2-Komponenten-System durch Phosphorylierungs-Signalisierungsmechanismen 19 . VirA / G aktiviert dann das gesamte vir- regulon, was zur Übertragung und Integration eines ~ 20 kb bakteriellen DNA-Fragments namens Transfer-DNA (T-DNA) aus seinem tumorinduzierenden (Ti) -Plasmid in den Pflanzenkern 16 führt . T-DNA trägt Gene, die für die Synthese der Pflanzenhormone Indol-3-essigsäure (IAA) ( iaaM und iaaH ) und Cytokinin ( ipt ) verantwortlich sind und einmal in Pflanzenzellen exprimiert werden, werden große Mengen dieser Phytohormone produziert. Dies führt zu einer abnormen Gewebevermehrung und Pflanzenentumorentwicklung, bekannt als Krone Gallenkrankheit, die ein chronisches und wiederauflebendes Problem für Pflanzen 9 , 11 , 20 ist . IAA wirkt auch gemeinsam mit Salicylsäure und Gamma-Aminobuttersäure, um Agrobacterium- Virulenz zu unterdrücken oder Agrobacteriu zu reduzieren M Quorum Sensing (QS) 17 , 21 , 22 . Um dieser Repression entgegenzuwirken, trägt T-DNA auch Gene für die Opin-Biosynthese, die Agrobacterium quorum-Sensing aktiviert, um die Agrobacterium- Pathogenität zu fördern und dient auch als Nährstoffquelle für den Pathogen 22 , 23 .
Trotz eines insgesamt tiefen Verständnisses von Agrobacterium- Pflanzen-Wechselwirkungen und der resultierenden T-DNA-Übertragung in den Pflanzenwirt sind die komplexen Signalisierungsereignisse im Anfangsstadium der Interaktion weniger gut verstanden. Dies ist teilweise auf die Einschränkungen konventioneller Ansätze zur Untersuchung von Agrobacterium- Plant-Signalisierung zurückzuführen. Pflanzenzell-Suspensionskulturen und künstliche ortsspezifische Verwundungen werden häufig verwendet, um molekulare Pflanzen-Mikroben-Wechselwirkungen zu untersuchen 24 ,Ef "> 26 , 27. Allerdings fehlen Zellsuspensionen einer typischen Pflanzenmorphologie, insbesondere haben Pflanzen-Suspensionszellen keine Wurzelstrukturen und Wurzel-Exsudate, die für die Aktivierung der mikrobiellen Chemotaxis und der Virulenz 28 , 29 sehr wichtig sind. Die Aufrechterhaltung der Pflanzenmorphologie Und die Wurzelstruktur wurde durch künstlich verwundete Pflanzen adressiert, was eine ortsspezifische Infektion erleichtert, was zum Nachweis von induzierten pflanzenverteidigungsbezogenen Genen in direkt infiziertem Pflanzengewebe 30 , 31 führt . Die künstliche Verwundung unterscheidet sich jedoch deutlich von der pathogenen Infektion in der Natur , Insbesondere wenn die Verwundung zu einer Jasmoninsäure (JA) -Akkumulation führt, die systemisch mit der natürlichen Anlagensignalisierung und -verteidigung in Einklang steht 26. Darüber hinaus werden synthetische Chemikalien typischerweise verwendet, um Pflanzenwirtsreaktionen künstlich zu induzierenOder pathogene Virulenz. Obwohl die Ergänzung solcher chemischen Verbindungen, die die Konzentrationen in Planta reflektieren, möglich ist, ist eine solche Ergänzung nicht die Diffusion von Wurzelausscheidungen allmählich in die umgebende Rhizosphäre, die einen von den Mikroben 28 , 32 erfaßten chemotaktischen Gradienten erzeugt. Angesichts der Einschränkungen herkömmlicher Ansätze zur Untersuchung von Pflanzen-Mikroben-Wechselwirkungen können die Genauigkeit und Tiefe der erhaltenen Daten behindert und restriktiv sein und das aus den konventionellen Ansätzen erzeugte Wissen kann nicht direkt in Planta umgesetzt werden . Viele Aspekte der Pflanzen- Agrobacterium- Signalisierung sind noch nicht vollständig verstanden, besonders in der frühen Phase der Wechselwirkungen, wenn die Krankheitssymptome noch nicht entwickelt haben.
Um die Grenzen herkömmlicher Ansätze zu ändern, stellt diese Arbeit eine kostengünstige, streng kontrollierbare und flexible Hydroponik dar. COkultivierungssystem, das es Forschern ermöglicht, tiefer Einblicke in die komplexen Signalisierungs- und Reaktionswege im Anfangsstadium molekularer Pflanzen-Mikroben-Wechselwirkungen zu gewinnen. Hydroponik wurde weithin verwendet, um Pflanzennährstoffe, Wurzel-Exsudate, Wachstumsbedingungen und die Auswirkungen der metallischen Toxizität auf Pflanzen 33 , 34 zu studieren. Es gibt mehrere Vorteile von hydroponischen Modellen, darunter die kleinen räumlichen Anforderungen, die Zugänglichkeit verschiedener Pflanzengewebe, die enge Kontrolle der Nährstoff- / Umweltbedingungen und die Schädlingsbekämpfung. Hydroponische Systeme sind auch weniger begrenzt auf Pflanzenwachstum im Vergleich zu Agar / Phytoagar-Beschichtung Techniken, die in der Regel beschränken Wachstum nach 2-3 Wochen. Wichtig ist, dass die Aufrechterhaltung von Ganzpflanzenstrukturen die für die mikrobielle Chemotaxis und die Virulenzinduktion notwendige natürliche Wurzelsekretion erleichtert 8 , 29 . Das System beschreibtBett hier ist einfacher und weniger arbeitsintensiv als die Alternativen 33 , 34 . Es nutzt weniger Teile und benötigt keine anderen Werkzeuge als Standardscheren. Es verwendet Metallgewebe (im Gegensatz zu Nylon 33 ) als eine starke Unterstützung für Pflanzenwachstum und eine einfache Methode der Belüftung unter sterilen Bedingungen durch Schütteln, um mikrobielles Wachstum zu unterstützen. Darüber hinaus kann das System Metallgewebe in verschiedenen Größen verwenden, um das Pflanzenwachstum zu unterstützen, das verschiedene Pflanzenarten aufnimmt, ohne die Breite ihrer Wurzeln zu beschränken.
In dem hier vorgestellten Hydroponik-Kokultur-System werden Pflanzen in einem sterilen Hydroponik-System angebaut, wo die Pflanzenwurzeln organische Verbindungen absondern, die das Wachstum von geimpften Bakterien unterstützen. In diesem Kokultivierungssystem werden keine künstlichen Chemikalien wie Pflanzenhormone, Verteidigungsauslöser oder virulenzinduzierende Chemikalien ergänzt, was die natürliche Zelle widerspiegelt-signale Homöostase bei Pflanzen-Mikroben-Wechselwirkungen. Mit diesem hydroponischen Kokultivierungssystem war es möglich, die Genexpression in Arabidopsis thaliana Col-0 Wurzelgewebe bei der Infektion durch Agrobacterium sowie die Aktivierung von Agrobacterium- Genen bei der Kokultivierung mit Arabidopsis gleichzeitig zu bestimmen. Es wurde weiter gezeigt, dass dieses System geeignet ist, Agrobacterium- Anlagerung an Pflanzenwurzeln sowie das Pflanzenwurzel-Sekretariat-Profil, bei der Kokultivierung (Infektion) mit Agrobacterium ( Abbildung 1 ) zu untersuchen.
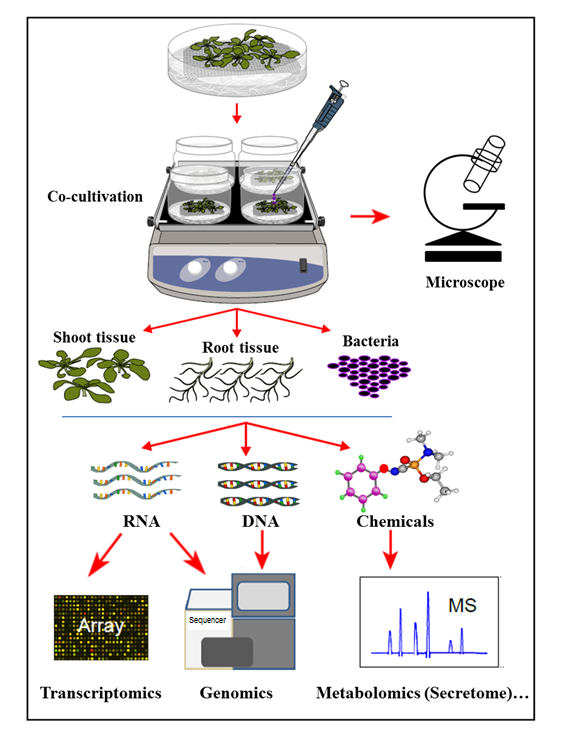
Abbildung 1: Überblick über das Hydroponic Cocultivation System mit Probenanalysen. Pflanzen werden oben auf dem Netz gezüchtet (schießt über dem Netz), wobei die Wurzeln in das hydroponische Medium eintauchen, das dann mit Bakterien geimpft wirdOder Kokultur. Pflanzengewebe und Bakterien werden dann für gleichzeitige Extraktionen und Analysen getrennt. Diese Zahl wurde aus Referenz 35 geändert.
