Planteassosierte mikrober spiller viktige roller i biogeokjemisk sykling, bioremediering, reduksjon av klimaendringer, plantevekst og helse og plantetoleranse mot biotiske og abiotiske påkjenninger. Mikroorganismer interagerer med planter både direkte gjennom plantecelleveggkontakt og indirekte via kjemisk sekresjon og signalering 1 , 2 , 3 . Som sessile organismer har planter utviklet direkte og indirekte mekanismer for å motstå infeksjon av patogener. Direkte forsvar inkluderer strukturelle forsvar og uttrykk for forsvarsproteiner, mens indirekte forsvar inkluderer sekundær plantemetabolittproduksjon og tiltrengning av organismer som er antagonistiske mot invaderende patogener 4 , 5 . Planteavledede rotutsprutninger, sekreter, slimhinner, mucigel og lysater forandrer de fysisk-kjemiske egenskapene til rhizosfæren for å tiltrekke seg eller frastøteMikrober mot sine verter 6 . Den kjemiske sammensetningen av rotsekresjon er artespesifikk, og derved tjener som et selektivt filter som tillater visse mikroorganismer som er i stand til å gjenkjenne slike forbindelser for å blomstre i rhizosfæren 6 . Således kan kompatible mikrobielle arter stimuleres til å aktivere og forbedre deres foreninger, enten til fordel for eller skade for planteverten 1 .
Forståelse av plante-mikrobe-interaksjoner i rhizosfæren er nøkkelen til å øke planteproduktiviteten og økosystemfunksjonen, siden et flertall av den mikrobielle og kjemiske eksponeringen forekommer ved rotstrukturen og jord-luftgrensesnittet 2 , 6 , 7 , 8 . Imidlertid har undersøkelsen av underjordiske plante-mikrobe-interaksjoner og gjensidige responser vært en utfordring på grunn av dens intrigerende Kompleks og dynamisk natur og mangel på egnede eksperimentelle modeller med naturlig rotstruktur og plantemorfologi under tett kontrollerbare vekstforhold. Som en av de mest studerte fytopathogenene, smitter Agrobacterium et bredt spekter av planter med landbruk og hagebruk, inkludert kirsebær, eple, pære, drue og rose 9 . Agrobacterium er en viktig modellorganisme for forståelse av plante-patogen-interaksjoner og er et kraftig verktøy i plantetransformasjon og anleggsteknologi 10 , 11 , 12 , 13 , 14 .
Molekylær plante- Agrobacterium- interaksjoner har blitt godt studert i flere tiår, og den nåværende forståelsen av Agrobacterium- patogenitet er omfattende 9 ,F.> 11 , 15 , 16. Agrobacterium- patogenitet er i stor grad tilskrevet sin evolusjonerte evne til å oppleve planteavledede signaler, noe som resulterer i finmoduleringen av dets virulensprogram og celle-til-cellekommunikasjon, såkalt quorum sensing 17 . Agrobacterium virulence-programmet er regulert av flere signaler tilgjengelig i rhizosfæren og involverer to sett med 2-komponentsystemer, ChvG / I-systemet og VirA / G-systemet. Syriske forhold i rhizosfæren aktiverer transkripsjonen av chvG / I , virA / G , Og flere andre gener involvert i Agrobacterium- patogenitet, inkludert virE0 , virE1 , virH1 , virH2 og gener av type VI-sekresjonssystemet (T6SS) 18. Plantavledede fenolforbindelser, inkludert acetosyringon (4'-hydroksy-3 ', 5 '-dimetoksyacetofenon), aktiver VIrA / G 2-komponentsystem gjennom fosforyleringssignalmekanismer 19 . VirA / G aktiverer så hele vir- regulonet, noe som resulterer i overføring og integrasjon av et 20 kb bakterielt DNA-fragment kalt overførings-DNA (T-DNA) fra sitt tumorinducerende (Ti) plasmid inn i plantekjernen 16 . T-DNA bærer gener ansvarlig for syntesen av plantehormonene indol-3-eddiksyre (IAA) ( iaaM og iaaH ) og cytokinin ( ipt ), og en gang uttrykt i planteceller, produseres store mengder av disse fytohormonene. Dette resulterer i unormal vevsproliferasjon og vekst i plantetumor, kjent som kronegangsykdom, som er et kronisk og resurgent problem for planter 9 , 11 , 20 . IAA virker også kollektivt med salisylsyre og gamma-amino smørsyre for å undertrykke Agrobacterium virulens eller å redusere Agrobacteriu M quorum sensing (QS) 17 , 21 , 22 . For å motvirke denne undertrykkelsen, bærer T-DNA også gener for opinbiosyntese, som aktiverer Agrobacterium quorum sensing for å fremme Agrobacterium- patogenitet og tjener også som næringsmiddelkilde for patogenet 22 , 23 .
Til tross for en generell dyp forståelse av Agrobacterium- planteinteraksjoner og den resulterende T-DNA-overføring til planteverten, er de komplekse signaleringshendelsene i den første fase av interaksjon mindre forstått. Dette skyldes delvis begrensningene av konvensjonelle tilnærminger for å undersøke Agrobacterium- plantesignalering. Plantecellesuspensjonskulturer og kunstig stedsspesifikke sår blir ofte brukt til å studere molekylære plante-mikrobe-interaksjoner 24 ,Ef "> 26 , 27. Imidlertid har cellesuspensjonene mangel på typisk plantemorfologi, spesielt plantesusjonsceller har ikke rotstrukturer og rotutsprutninger, noe som er svært viktig for aktivering av mikrobiell kjemotaks og virulens 28 , 29. Vedlikehold av plantemorfologi Og rotstruktur har blitt tatt opp av kunstig sårende planter, noe som letter tilstedsspesifikke infeksjoner, noe som resulterer i deteksjon av induserte planteforsvarrelaterte gener i direkte infisert plantevev 30 , 31. Imidlertid er kunstig sår betydelig forskjellig fra patogeninfeksjon i naturen , Særlig ettersom sår fører til akkumulering av jasmonisk syre (JA), som systematisk forstyrrer naturlig plantesignalering og -forsvar 26. I tillegg er syntetiske kjemikalier vanligvis brukt til kunstig framkalling av plantevertsresponserEller patogen virulens. Selv om tilskudd av slike kjemiske forbindelser som reflekterer konsentrasjoner i planta, er mulig, utgjør slik tillegg ikke diffusjonen av rotutsprutninger gradvis inn i den omkringliggende rhizosfæren, noe som genererer en kjemotaktisk gradient registrert av mikrober 28 , 32 . Gitt begrensningene ved konvensjonelle tilnærminger for å studere plante-mikrobe-interaksjoner, kan nøyaktigheten og dybden av de innhentede dataene bli hindret og restriktiv, og kunnskapen generert fra de konvensjonelle tilnærmingene kan ikke oversettes direkte i planta . Mange aspekter av plante- Agrobacterium- signalering er ennå ikke fullt ut forstått, særlig i det tidlige stadiet av interaksjoner, når sykdomssymptomene ennå ikke er utviklet.
For å endre grensene for konvensjonelle tilnærminger, presenterer dette arbeidet en billig, tett kontrollerbar og fleksibel hydroponisk cOdlingssystem som gjør det mulig for forskere å få dypere innblikk i de komplekse signal- og responsveiene ved første fase av molekylære plante-mikrobe-interaksjoner. Hydroponics har vært mye brukt til å studere plantenæringsstoffer, rotutsprutninger, vekstbetingelser og virkningen av metallisk toksisitet på planter 33 , 34 . Det finnes flere fordeler med hydroponiske modeller, inkludert de små romlige kravene, tilgjengeligheten til forskjellige plantevev, den tette kontrollen av næringsstoffer / miljøforhold og bekjempelse av skadedyr / sykdom. Hydroponiske systemer er også mindre begrensende for plantevekst i forhold til agar / phytoagar plating teknikker, som vanligvis begrenser veksten etter 2-3 uker. Viktig er at vedlikehold av helplantestrukturer letter den naturlige rotsekresjonen som er nødvendig for mikrobiell kjemotaks og virulensinduksjon 8 , 29 . SystembeskrivelsenSeng her er enklere og mindre arbeidsintensiv enn alternativene 33 , 34 . Den bruker færre deler og krever ikke andre verktøy enn standard saks. Den bruker metallnett (i motsetning til nylon 33 ) som en sterk støtte for plantevekst og en enkel metode for lufting under sterile forhold gjennom risting for å støtte mikrobiel vekst. I tillegg kan systemet bruke metallnett av forskjellige størrelser for å støtte plantevekst, som imøtekommer mangfoldige plantearter uten å begrense bredden på sine røtter.
I det hydroponiske kultiveringssystemet som presenteres her, dyrkes planter i et sterilt hydroponisk system hvor plantenavnene utskiller organiske forbindelser som støtter veksten av inokulerte bakterier. I dette kultiveringssystemet blir ingen kunstige kjemikalier, som plantehormoner, forsvarselektronik eller virulensinducerende kjemikalier, supplert, noe som reflekterer den naturlige cellenSignalerende homeostase under plante-mikrobe interaksjoner. Med dette hydroponiske kultiveringssystemet var det mulig å samtidig bestemme genuttrykk i Arabidopsis thaliana Col-0 rotvev på infeksjon av Agrobacterium , samt aktivering av Agrobacterium- gener ved kultivering med Arabidopsis . Det ble videre påvist at dette systemet er egnet til å studere Agrobacterium- vedlegg til planterøtter, så vel som planterotsekretomprofilen, ved kultivering (infeksjon) med Agrobacterium ( Figur 1 ).
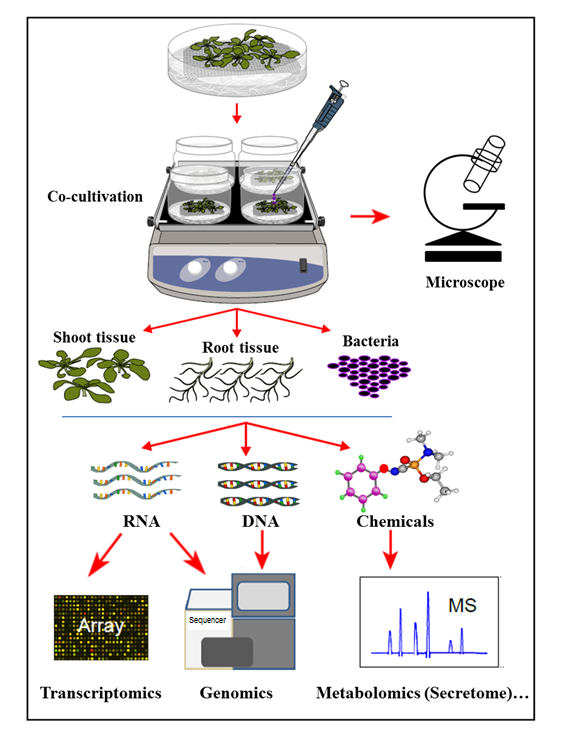
Figur 1: Oversikt over det hydroponiske kultiveringssystemet, med prøveanalyser. Planter dyrkes på toppen av masken (skudd over masken), med røttene nedsenket i hydroponisk medium som deretter inokuleres med bakterier fEller kultur. Plantevev og bakterier separeres deretter for samtidige ekstraksjoner og analyser. Denne figuren er endret fra referanse 35 .
