Växtassocierade mikrober spelar viktiga roller inom biogeokemisk cykling, bioremediering, minskning av klimatförändringar, växttillväxt och hälsa och växt tolerans mot biotiska och abiotiska påfrestningar. Mikroorganismer interagerar med växter både direkt genom växtcellsväggkontakt och indirekt via kemisk utsöndring och signalering 1 , 2 , 3 . Som sessila organismer har växter utvecklat direkta och indirekta mekanismer för att motstå infektion med patogener. Direkta försvar inkluderar strukturella försvar och uttryck av försvarproteiner, medan indirekta försvar inkluderar sekundär växtmetabolitproduktion och attraktion av organismer som är antagonistiska mot invaderande patogener 4 , 5 . Växthärdiga rotutsprutningar, sekret, mukilager, mucigel och lysater förändrar rhizosfärens fysikalisk-kemiska egenskaper för att locka till sig eller avvisaMikrober mot sina värdar 6 . Den kemiska sammansättningen av rotsekretion är artsspecifik och tjänar därigenom som ett selektivt filter som tillåter vissa mikroorganismer som kan känna igen sådana föreningar för att blomstra i rhizosfären 6 . Således kan kompatibla mikrobiella arter stimuleras för att aktivera och förbättra deras föreningar, antingen till fördel eller nackdel för växtvärden 1 .
Förstå växelmikroskopiska interaktioner i rhizosfären är nyckeln till att förbättra växtproduktiviteten och ekosystemets funktion, eftersom en majoritet av den mikrobiella och kemiska exponeringen förekommer vid rotstrukturen och markluftsgränssnittet 2 , 6 , 7 , 8 . Undersökningen av underjordiska växt-mikrobi-interaktioner och ömsesidiga svar har dock varit en utmaning på grund av dess intrigerande Komplex och dynamisk natur och bristen på lämpliga experimentella modeller med naturlig rotstruktur och växtmorfologi under tätt kontrollerbara tillväxtförhållanden. Som ett av de mest studerade fytopathogenerna infekterar Agrobacterium ett brett sortiment av växter med jordbruks- och trädgårdsbeteende, inklusive körsbär, äpple, päron, druva och ros 9 . Agrobacterium är en viktig modellorganisme för förståelse av växtpatogen-interaktioner och är ett kraftfullt verktyg vid växtomvandling och växtteknik 10 , 11 , 12 , 13 , 14 .
Molekylära växt- Agrobacterium- interaktioner har studerats väl i flera årtionden, och den nuvarande förståelsen av Agrobacterium- patogeniciteten är omfattande 9 ,F "> 11 , 15 , 16. Agrobacteriumpatogeniteten är till stor del hänförlig till dess utvecklade förmåga att uppleva växtgenererade signaler, vilket resulterar i finmoduleringen av dess virulensprogram och cell-till-cell-kommunikation, så kallad kvorumavkänning 17 . Agrobacterium virulence-programmet regleras av flera signaler som finns tillgängliga i rhizosfären och innefattar två uppsättningar av 2-komponentsystem, ChvG / I-systemet och VirA / G-systemet. Syrabetingelser i rhizosfären aktiverar transkriptionen av chvG / I , virA / G , Och flera andra gener som är involverade i Agrobacterium- patogenitet, inklusive virE0 , virE1 , virH1 , virH2 och gener av typ VI-sekretionssystemet (T6SS) 18. Växtgenererade fenoliska föreningar, innefattande acetosyringon (4'-hydroxi-3 ', 5 '-dimetoxiacetofenon), aktivera VIrA / G 2-komponentsystem genom fosforyleringssignalmekanismer 19 . VirA / G aktiverar sedan hela vir- regulon, vilket resulterar i överföring och integration av ett ~ 20 kb bakteriellt DNA-fragment som kallas överförings-DNA (T-DNA) från sin tumörinducerande (Ti) plasmid i växtkärnan 16 . T-DNA bär gener ansvarig för syntesen av växthormonerna indol-3-ättiksyra (IAA) ( iaaM och iaaH ) och cytokinin ( ipt ) och en gång uttryckt i växtceller produceras stora mängder av dessa fytohormoner. Detta resulterar i onormal vävnadsproliferation och utveckling av växttumörer, känd som krongallsjukdom, vilket är ett kroniskt och återkommande problem för växter 9 , 11 , 20 . IAA verkar också gemensamt med salicylsyra och gamma-amino-smörsyra för att undertrycka Agrobacterium virulens eller för att minska Agrobacteriu M kvorumavkänning (QS) 17 , 21 , 22 . För att motverka denna repression, bär T-DNA också gener för opinbiosyntes, vilken aktiverar agrobacteriumkvumumavkännande för att främja Agrobacterium- patogenicitet och tjänar också som en näringskälla för patogenen 22 , 23 .
Trots en övergripande djup förståelse för Agrobacterium- plant interaktioner och den resulterande T-DNA-överföringen i växtvärden är de komplexa signaleringshändelserna vid det inledande skedet av interaktion mindre väl förstådda. Detta beror delvis på begränsningarna av konventionella tillvägagångssätt för att undersöka Agrobacterium- plantsignalering. Växtcellssuspensionskulturer och artificiell platsspecifik sårning används vanligen för att studera molekylära växt-mikrobe-interaktioner 24 ,Ef "> 26 , 27. Cellsuspensioner saknar typiskt växtmorfologi, i synnerhet har plantesuspensionsceller inte rotstrukturer och rotutsöndringar, vilka är mycket viktiga för aktivering av mikrobiell kemotax och virulens 28 , 29. Behållandet av växtmorfologi Och rotstruktur har behandlats av artificiellt sårande växter, vilket underlättar platsspecifik infektion, vilket resulterar i detektion av inducerade växtförsvarsrelaterade gener i direkt infekterad vävnad 30 , 31. Men artificiell sårning skiljer sig signifikant från patogeninfektion i naturen , Speciellt som att sår leder till ackumulering av jasmonsyra (JA), som systemiskt stör naturligt växtsignalering och försvar 26. Dessutom används syntetiska kemikalier typiskt för att artificiellt inducera växtvärdesreaktionerEller patogen virulens. Även om tillägget av sådana kemiska föreningar som reflekterar koncentrationer i plantan är möjligt, svarar sådant tillskott inte för diffusion av rotutsprutning gradvis i den omgivande rhizosfären, vilken alstrar en kemotaktisk gradient som avkännes av mikrober 28 , 32 . Med tanke på begränsningarna av konventionella metoder för att studera växelmikroskopiska interaktioner kan noggrannheten och djupet hos de erhållna data hindras och begränsas, och kunskapen som genereras av konventionella metoder kan inte översättas direkt i plantan . Många aspekter av växt- Agrobacterium- signaleringen är ännu inte helt förstådda, särskilt i det tidiga skedet av interaktioner, när sjukdomssymptomerna ännu inte har utvecklats.
För att ändra gränserna för konventionella tillvägagångssätt presenterar detta arbete en billig, tätt kontrollerbar och flexibel hydroponisk cOdlingssystem som gör det möjligt för forskare att få djupare insikter i de komplexa signal- och reaktionsvägarna vid det inledande skedet av molekylära växtmikro-interaktioner. Hydroponics har använts i stor utsträckning för att studera växtnäringsämnen, rotutsöndringar, tillväxtförhållanden och effekterna av metallisk toxicitet på växter 33 , 34 . Det finns flera fördelar med hydroponiska modeller, inklusive de små rumsliga kraven, tillgången till olika vävnader, den täta kontrollen av näringsämnen / miljöförhållandena och skadedjursbekämpningen. Hydroponiska system är också mindre begränsande för växttillväxt i jämförelse med agar / fytoagarpläteringstekniker, som typiskt begränsar tillväxten efter 2-3 veckor. Viktigt är att underhållet av helplanta strukturer underlättar den naturliga rotsekretionen som är nödvändig för mikrobiell kemotax och virulensinduktion 8 , 29 . Systemet beskrivsSängen här är enklare och mindre arbetsintensiv än alternativen 33 , 34 . Det använder färre delar och kräver inga andra verktyg än standard sax. Det använder metallnät (i motsats till nylon 33 ) som ett starkt stöd för växttillväxt och en enkel metod för luftning under sterila betingelser genom att skaka för att stödja mikrobiell tillväxt. Dessutom kan systemet använda metallnät av olika storlekar för att stödja växttillväxt, vilket rymmer olika växtarter utan att begränsa bredden på sina rötter.
I det hydroponiska odlingssystemet som presenteras här odlas växter i ett sterilt hydroponiskt system där växtrötterna utsöndrar organiska föreningar som stöder tillväxten av inokulerade bakterier. I detta odlingssystem kompletteras inga konstgjorda kemikalier, såsom växthormoner, försvar elicitor eller virulence-inducerande kemikalier, vilket återspeglar den naturliga cellenSignalerande homeostas under interaktioner mellan växter och mikroorganismer. Med detta hydroponiska kultiveringssystem var det möjligt att samtidigt bestämma genuttryck i Arabidopsis thaliana Col-0-vävnad vid infektion med Agrobacterium , såväl som aktiveringen av Agrobacterium- gener vid odling med Arabidopsis . Det visades vidare att detta system är lämpligt att studera Agrobacterium- bindning till växtrotsar, liksom växtrodssekretomprofilen, vid odling (infektion) med Agrobacterium ( Figur 1 ).
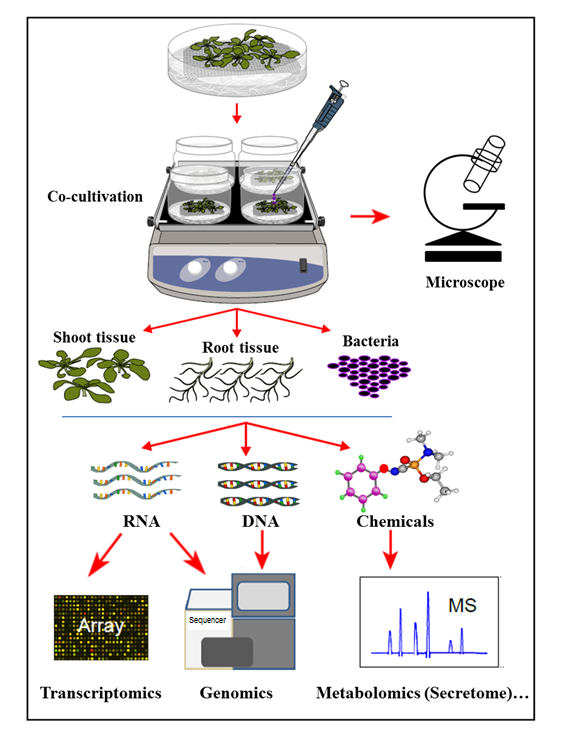
Figur 1: Översikt över det hydroponiska kultiveringssystemet, med provanalyser. Växter odlas på nätet (skott över nätet), med rötterna nedsänkta i hydroponiskt medium som sedan inokuleras med bakterier fEller odling. Växtvävnader och bakterier separeras sedan för samtidiga extraktioner och analyser. Denna siffra har ändrats från referens 35 .
