بعض أنواع الأكسجين التفاعلية (ROS) قادره علي أكسده الروابط المزدوجة الكربون من قواعد الحمض النووي وبعض الكربونات في moiety ديوكسيريبوز ، وتوليد القواعد المؤكسدة وفواصل حبلا الحمض النووي1. كجزيء مشحونة سلبا الغنية في ذرات النيتروجين والأكسجين ، والحمض النووي هو أيضا هدف للمجموعات الكهربائية التي تتفاعل بشكل تساهميا مع المواقع النونويه (النيتروجين والأكسجين) ، وإعطاء المنتجات التي تسمي الحمض النووي القناات2. لذلك ، الحمض النووي الأنابيب والأحماض النووية المؤكسدة هي أمثله من آفات الحمض النووي التي هي مفيده العلامات الحيوية لتقييم السمية من المواد التي هي الكتروفيريك ، توليد الكهربائية رد الفعل علي التحول الإحيائي ، أو الحث علي الاكسده1، 2– علي الرغم من ان قواعد الحمض النووي المعدلة يمكن ازالتها من الحمض النووي عن طريق إصلاح الاستئصال القاعدي أو النوكليوتيد (BER أو NER) ، فان التحريض علي عدم التوازن بين توليد وأزاله آفات الحمض النووي لصالح السابق يؤدي إلى زيادة صافيه في مستوياتها في العمل الإضافي للحمض النووي3 . النتائج هي زيادة معدلات طفرة الحمض النووي ، وانخفاض التعبير الجيني ، وتقلص النشاط البروتين2،4،5،6،7، والآثار التي ترتبط ارتباطا وثيقا تطور الامراض. قد تؤثر طفرات الحمض النووي علي وظائف خلوية متنوعة ، مثل إشارات الخلايا ، ودوره الخلايا ، وسلامه الجينوم ، واستقرار التيتيمير ، والنعوت ، وهيكل الكروماتين ، والربط الريبي ، وتوازن البروتين ، والأيض ، والخلايا المبرمجة ، وتمايز الخلية8 ،9. استراتيجيات لإبطاء معدلات الطفرة الخلية وتطوير الامراض المزمنة (مثل السرطان, امراض الأعصاب) تمر من خلال معرفه مصادر الطفرة, من بينها, آفات الحمض النووي وأسبابها.
ومن الأسباب الهامه للضرر الجزيء ، بما في ذلك الحمض النووي وتلف الدهون ، الاصابه بالامراض المزمنة بسبب التعرض للتلوث ، والتهابالمستمر ، والمرض الباثولوجي المرضي (مثل مرض السكري) ، وما إلى ذلك. وعلي سبيل المثال ، فان الراديكالية الهيدروكسيل عاليه التفاعل (OH) شكلت من H2O2 الحد من خلال الانتقال أيونات المعادن (Fe2 +، Cu +) أكسده قواعد الحمض النووي ، والحمض النووي السكر شارده والأحماض الدهنية غير المشبعة في السيطرة علي انتشار معدلات10. من بين 80 التي تتميز بالفعل النيوبواسات المؤكسدة3، والأكثر دراسة واحد هو 8-oxo-7 ، 8-ديهيدروجوانيني (8-oxoGua) أو 8-oxo-7 ، 8-ديهيدرو-2’غوانوسين (8-oxodGuo ، الشكل 1) ، وهي آلافه التي هي قادره علي الحث GT transversions خلايا الثدييات10،11. يتم تشكيلها من قبل أحاديه الاكسده الكترونيه من الغوانين ، أو بواسطة الهيدروكسيل الراديكالية أو الهجوم الأكسجين قميص من الغوانين في الحمض النووي1. الأحماض الدهنية غير المشبعة هي الأهداف الهامه الأخرى للاكسده عاليه التفاعل, مثل •آوه, التي تبدا عمليه الاكسده الدهنية1,12. فانه يعطي ارتفاعا إلى أكاسيد الهيدروجين الأحماض الدهنية التي قد تتحلل لالالديهيدات الكهربائية و ابكسييلديهيديس, مثل مالونوديهايد, 4-هيدروكسي-2-نونينال, 2, 4-ديكادينال, 4, 5-الايبوكسي-(2ه)-ديسينال, hexenal, اكرولاين, كروتونالديهيد, وهي قادره علي تشكيل مغيرات الحمض النووي المبيدات الوراثية ، مثل مالونوديهايد-، propano-، أو etheno ادوكتس1،12،13. القناات etheno 1,n2-etheno-2 ‘-غوانوسين (1,n2-εdguo, الشكل 1) و 1,n6-ايثينو-2 ‘-ديوكسينونوسين (1,ن6-εdado, الشكل 1 ) وقد اقترح كالمؤشرات الحيوية المحتملة في الفيزيولوجيا المرضية للالتهاب14،15.
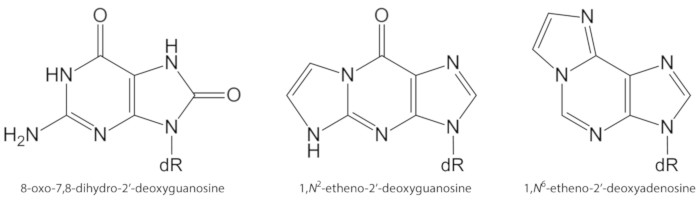
الشكل 1 الهياكل الكيميائية للآفات الحمض النووي الكمية في هذه الدراسة. الدكتور = 2 ‘ ديوكسيريبوز. وقد تم تعديل هذا الرقم من اوليفيرا وآخرون34. يرجى النقر هنا لعرض نسخه أكبر من هذا الرقم.
سمحت الدراسات التي أجريت في أوائل الثمانينات بالكشف الحساس ل 8-oxodGuo بواسطة اللوني السائل عالي الأداء إلى جانب الكشف الكهروكيميائي (HPLC-نماء الطفولة المبكرة). القياس الكمي من 8-oxodguo بواسطة hplc-النماء في الطفولة الباكرة في العديد من النظم البيولوجية التي خضعت لظروف مؤكسده أدت إلى الاعتراف 8-oxodguo كمؤشر حيوي من الضرر قاعده المستحثة بالاكسده في الحمض النووي1,16. علي الرغم من ان قويه والسماح للقياس الكمي من 8-oxodGuo في نطاق fmol منخفضه17, قياسات hplc-النماء في مرحله الطفولة المتوسطة تعتمد علي دقه الوقت الاحتفاظ التحليلية لتحديد التحليل وعلي القرار اللوني لتجنب التدخلات من مكونات عينه أخرى. كما يتطلب الكشف الكهروكيميائية استخدام الملح (علي سبيل المثال ، فوسفات البوتاسيوم ، خلات الصوديوم) في المرحلة المتنقلة ، والحفاظ علي الظروف التحليلية المناسبة يحتاج العمود الروتيني والمعدات وقت التنظيف.
بدلا من ذلك ، فان استخدام الحمض النووي البكتيرية إصلاح الانزيم الحمض النووي الجليكوسيلاسي (fpg) و ، بعد ذلك ، الإنسان 8-oxoguanine جليكوسيلاسي 1 (hOGG1) ، للكشف عن وأزاله 8-oxogua من الحمض النووي ، برزت كوسيلة لتحريض الحمض النووي القلوي عطوب مواقع. يتم تحويل المواقع القلوية عطوب إلى فواصل حبلا الحمض النووي والسماح للقياس الدقيق غير المباشر عاليه جدا من 8-oxogua القلوية خليه واحده هلام الكهربائي (“فحص المذنب”). الحساسية العالية وإنجاز التحاليل دون الحاجة إلى استخراج الحمض النووي الخلوي هي المزايا الرئيسية لهذا النوع من الفحص. فانه يعطي ادني مستويات الحالة الثابتة من 8-oxoGua في الحمض النووي ، عاده 7-10 مرات اقل من المستويات التي تم الحصول عليها من قبل الأساليب التحليلية الحيوية علي أساس HPLC. ومع ذلك ، فمن قياس غير مباشر من 8-oxogua وبعض العيوب هي عدم وجود خصوصية أو الكفاءة غير معروفه لانزيمات الإصلاح المستخدمة1،16،18.
اختبارات المناعية هي مجموعه أخرى من الأساليب المستخدمة للكشف عن 8-oxoGua1 و adducts الحمض النووي exocyclic, مثل 1,n6-dado و 1,n2-dado12. علي الرغم من الحساسية ، والقصور في استخدام الأجسام المضادة للكشف عن آفات الحمض النووي هو عدم وجود خصوصية بسبب التفاعل التبادلي إلى مكونات أخرى من العينات البيولوجية ، بما في ذلك قواعد الحمض النووي الطبيعي1،12. ويمكن أيضا الكشف عن الحمض النووي المبيدات ، بما في ذلك 1 ،n6-Dado و 1 ،n2-dado ، وتحديدها كميا من قبل 32حساسة للغاية المقايسات P-postlabeling12. حساسية عاليه من 32P-التسمية البريدية يسمح باستخدام كميات صغيره جدا من الحمض النووي (علي سبيل المثال ، 10 ميكروغرام) للكشف عن حوالي 1 معقد إضافي لكل 10 10 قواعد طبيعيه19. ومع ذلك ، فان استخدام المواد الكيميائية الراديويه ، وعدم وجود خصوصية كيميائية وانخفاض الدقة هي بعض العيوب19،20.
والتقييد المشترك للأساليب المذكورة أعلاه هو الانتقائية أو التحديد المنخفض للكشف عن الجزيئات المطلوبة. في هذا السيناريو ، HPLC بالاضافه إلى التاين بالكهرباء بالتوازي الطيف الكتلي (HPLC-ESI-MS/MS و HPLC-MS3) تطورت كمعيار الذهب للقياس الكمي من النيونوسيدس المعدلة في المصفوفات البيولوجية ، مثل الحمض النووي والبول والبلازما واللعاب 1 , 19 , 20. مزايا HPLC-ESI-MS/ms طرق هي حساسية (عاده في نطاق fmol منخفضه) والخصوصية العالية التي قدمتها ط) الانفصال الكروماتوغرافي ، الثاني) السمة والمعروفة نمط من تجزئه جزيء داخل كتله مطياف الاصطدام الغرفة ، والثالث) القياس الدقيق للكتلة المحددة لتهمه نسبه (m/z) في رد فعل متعددة وضع الرصد1،19. ويضيف استخدام المعايير الداخلية المسمية بالنظائر ميزه التصحيحات المتعلقة بخسائر الجزيئات اثناء التحلل بالحمض النووي والخطوات التحليلية للتحليل ، فضلا عن الاختلافات في التاين التحليلي بين العينات. هو أيضا [ايدس] في التحديد من ال يصح [كروغرفيك] قمة عندما أكثر من واحده قمة يكون حاضره1,12,19,20.
وقد استخدمت عده طرق علي أساس hplc-ESI-ms/ms للقياس الكمي من 8-oxodguo, 1,n6-dado و 1,n2-dado في الحمض النووي المستخرج من عينات بيولوجية مختلفه12,15,20 ،21،22،23،24،25،26،27،28،29 . الجسيمات الدقيقة (PM2.5) تحمل المواد الكيميائية العضوية وغير العضوية ، مثل الهيدروكربونات العطرية متعددة الدوريات ، نيترو-pahs ، الديهيديس ، كيتون ، والأحماض الكربوكسيليه ، الكينولين ، والمعادن ، والأيونات القابلة للذوبان في الماء ، والتي قد تحفز التهاب الاكسده ، والظروف التي تحبذ حدوث الضرر جزيء والمرض30،31،32،33. نقدم هنا التحقق من صحة HPLC-ESI-MS/MS الأساليب التي تم تطبيقها بنجاح للقياس الكمي من 8-oxodGuo ، 1 ،n6-Dado و 1 ،n2-dado في الرئة والكبد والكلي الحمض النووي من الفئران A/J لتقييم اثار المحيطة PM2.5 التعرض34.
