Algumas espécies reativas de oxigênio (ROS) são capazes de oxidar ligações duplas de carbono de bases de DNA e alguns carbonos na fração desoxirribose, gerando bases oxidadas e quebras de fio de DNA1. Como uma molécula carregada negativamente rica em átomos de nitrogênio e oxigênio, o DNA também é um alvo para grupos eletrofílicos que reagem covalentemente com os sítios nucleofílicos (nitrogênio e oxigênio), dando produtos que são chamados de adutos de DNA2. Assim, os adutos do ADN e as bases oxidadas do ADN são exemplos de lesões do ADN que são biomarcadores úteis para a avaliação da toxicidade das substâncias que são electrofílicas, geram eletrófilos reactivos em cima da biotransformação, ou induzem o stress oxidativo1, o 2. Embora as bases modificadas do ADN possam ser removidas do ADN pela base ou pelo reparo da excisão do nucleotide (BER ou NER), a indução de um desequilíbrio entre a geração e a remoção de lesões do ADN em favor do anterior conduz a um aumento líquido de seus níveis no tempo estipulado do ADN3 . Os resultados são o aumento das taxas de mutação do DNA, a redução da expressão gênica e a diminuição da atividade protéica2,4,5,6,7, efeitos que estão intimamente relacionados com a desenvolvimento de doenças. As mutações do DNA podem afetar diversas funções celulares, como sinalização celular, ciclo celular, integridade do genoma, estabilidade do telômero, epigenome, estrutura da cromatina, splicing de RNA, homeostase proteica, metabolismo, apoptose e diferenciação celular8 ,9. Estratégias para retardar as taxas de mutação celular e o desenvolvimento crônico da doença (por exemplo, câncer, doenças neurodegenerativas) passam pelo conhecimento das fontes de mutação, entre elas, as lesões do DNA e suas causas.
ROS gerados endogenamente em excesso, devido à exposição ao poluente, inflamação persistente, fisiopatologia da doença (por exemplo, diabetes), etc., são causas importantes de dano da biomolécula, incluindo o DNA e o dano lipídico1. Como exemplo, o radical hidroxila altamente reativo (OH) formado a partir de H2O2 redução por íons metálicos de transição (FE2 +,+ +) oxida as bases de DNA, a fração de açúcar de DNA e os ácidos graxos poliinsaturados em controle de difusão taxas de10. Entre os 80 já caracterizados nucleobases oxidadas3, o mais estudado é 8-oxo-7,8-diidroguanina (8-oxoGua) ou 8-oxo-7,8-dihidro-2 ‘-Desoxiguanosina (8-oxodGuo, Figura 1), uma lesão que é capaz de induzir transversões gt em células mamíferas10,11. É formado pela oxidação eletrônica mono da guanina, ou pelo ataque de oxigênio radical hidroxila ou singlet de guanina no DNA1. Os ácidos graxos poliinsaturados são outros alvos importantes de oxidantes altamente reativos, como •Oh, que iniciam o processo de peroxidação lipídica1,12. Ele dá origem a hidroperóxidos de ácidos graxos que podem se decompor a aldeídos eletrofílicos e epoxialdeídos, tais como malondialdeído, 4-hidroxi-2-nonenal, 2,4-decadienal, 4, 5-epóxi-(2E)-decenal, hexenal, acroleína, crotonaldeído, que são capaz de formar adutos de DNA exocílicos mutagénicos, como os adutos de malondialdeído, propano ou eteno1,12,13. Os adutos de eteno 1,n2-eteno-2′-deoxyguanosine (1,n2-εdguo , Figura 1) e 1,n6-eteno-2 ‘-deoxyadenosine (1,n6-εdado, Figura 1 ) têm sido sugeridas como potenciais biomarcadores na fisiopatologia da inflamação14,15.
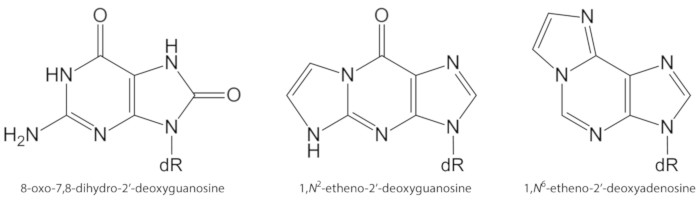
Figura 1. Estruturas químicas das lesões de DNA quantificadas no presente estudo. dR = 2 ́-desoxirribose. Este número foi modificado de Oliveira et al.34. Por favor clique aqui para ver uma versão maior desta figura.
Estudos realizados no início da década de 1980 permitiram a detecção sensível de 8-oxodGuo por cromatografia líquida de alta eficiência acoplada à detecção eletroquímica (HPLC-ECD). A quantificação de 8-oxodguo por HPLC-ECD em diversos sistemas biológicos sujeitados às circunstâncias oxidantes conduziu ao Recognition de 8-oxodguo como um biomarcador de dano de base oxidativamente induzido no ADN1,16. Embora robusto e permitindo a quantificação de 8-oxodguo na escala baixa de fmol17, as medidas do HPLC-ECD confiam na exatidão do tempo da retenção do analito para a identificação do analito e na definição da cromatografia para evitar interferências de outros constituintes da amostra. Como a detecção eletroquímica requer o uso de sal (por exemplo, fosfato de potássio, acetato de sódio) na fase móvel, a manutenção de condições analíticas adequadas precisa de rotina de coluna e tempo de limpeza do equipamento.
Alternativamente, o uso do DNA bacteriano da enzima de reparo do ADN glicosilase do formamidopyrimidine (fpg) e, mais tarde, o glicolisase 1 do humano 8-oxoguanine (hOGG1), para a deteção e a remoção de 8 oxogua do ADN, emergiu como uma maneira para a indução do lábil do alcalóide do ADN Sites. Os locais lábeis do alcalóide são convertidos às rupturas da costa do ADN e permitem a quantificação indireta muito elevada sensível de 8 oxoGua pela electroforese alcalina do gel da única pilha (“ensaio do cometa”). A alta sensibilidade e a realização das análises sem a necessidade de extração de DNA celular são as principais vantagens deste tipo de ensaio. Dá os níveis os mais baixos do estado estacionário de 8 oxoGua no ADN, tipicamente 7-10 vezes mais baixos do que os níveis obtidos por métodos bioanalíticos baseados no HPLC. No entanto, é uma medida indireta de 8-oxogua e algumas desvantagens são a falta de especificidade ou a eficiência desconhecida das enzimas de reparo utilizadas1,16,18.
Os imunoensaios são outro conjunto de métodos utilizados para a detecção de 8-oxoGua1 e ADUTOS de DNA exocídrico, como 1,n6-dAdo e 1,n2-dguo12. Apesar da sensibilidade, uma lacuna do uso de anticorpos para a detecção de lesões de DNA é a falta de especificidade devido à reatividade cruzada a outros componentes de amostras biológicas, incluindo as bases normais de DNA1,12. Os adutos exocílicos de DNA, incluindo 1, n6-DAdo e 1, n2-dguo, também podem ser detectados e quantificados por ensaios altamente sensíveis de 32P-postetiquetagem12. A sensibilidade elevada de 32P-postrotulagem permite o uso de quantidades muito pequenas de ADN (por exemplo, 10 μg) para a deteção de aproximadamente 1 aduto por 1010 bases normais19. No entanto, o uso de radioquímicos, a falta de especificidade química e a baixa acurácia são algumas desvantagens19,20.
Uma limitação compartilhada dos métodos citados acima é a baixa seletividade ou especificidade para a detecção das moléculas desejadas. Nesse cenário, a HPLC acoplada à espectrometria de massas em tandem de ionização por electrospray (HPLC-ESI-MS/MS e HPLC-MS3) evoluiu como padrão-ouro para quantificação de nucleosídeos modificados em matrizes biológicas, como DNA, urina, plasma e saliva 1. º , 19 anos de , 20. vantagens dos métodos de HPLC-ESI-MS/MS são a sensibilidade (tipicamente na baixa escala do fmol) e a especificidade elevada fornecida por i) a separação cromatográfica, II) o teste padrão característico e conhecido da fragmentação da molécula dentro da massa Câmara de colisão do espectrómetro, e III) a medida exata da massa selecionada para carregar a relação (m/z) no modo múltiplo da monitoração da reação1,19. O uso de padrões internos isotopicamente rotulados acrescenta a vantagem de correções para perdas de moléculas durante a hidrólise do DNA e as etapas de enriquecimento do analito, bem como para as diferenças da ionização do analito entre as amostras. Auxilia também na identificação do pico cromatográfico correto quando mais de um pico está presente1,12,19,20.
Vários métodos baseados em HPLC-ESI-MS/MS têm sido utilizados para quantificação de 8-oxodguo, 1,n6-dAdo e 1,n2-dguo em DNA extraído de diferentes amostras biológicas12,15,20 ,21,22,23,24,25,26,27,28,29 . Partículas finas (PM2,5) transportar produtos químicos orgânicos e inorgânicos, tais como hidrocarbonetos policíclicos aromáticos (PAHS), Nitro-PAHS, aldeídos, cetonas, ácidos carboxílicos, quinolinas, metais e íons solúveis em água, que podem induzir inflamação e estresse oxidativo, condições que favorecem a ocorrência de danos biomoléculas e doença30,31,32,33. Nós apresentamos aqui métodos validados de HPLC-ESI-MS/MS que foram aplicados com sucesso para a quantificação de 8-oxodGuo, de 1, den6-dAdo e de 1, den2-dguo no ADN do pulmão, do fígado e do rim de ratos de a/J para a avaliação do efeitos do ambiente PM2,5 exposição34.
