כמה מיני חמצן תגובתי (ROS) הם מסוגלים החמצן איגרות חוב כפול של DNA בסיסים ו כמה פחמנים deoxyribose moiety, יצירת בסיסים חמצון ו-DNA שובר הגדיל1. כמולקולה טעונה שלילית עשיר חנקן ואטומי חמצן, DNA הוא גם יעד עבור קבוצות electrophilic כי מגיבים בעקשנות עם האתרים הנופילים (חנקן וחמצן), מתן מוצרים הנקראים adducts DNA2. כך, adducts DNA ו תחמוצת בסיסים DNA הם דוגמאות של נגעים DNA כי הם ביטויים שימושיים עבור הערכה רעילות של חומרים, כי הם electrophilic, ליצור אלקטרופילנים תגובתי על הטרנספורמציה ביולוגית, או לגרום למתח חמצוני1, 2. שתיים. למרות בסיסים DNA שונה ניתן להסיר דנ א על ידי בסיס או נוקלאוטיד תיקון כריתה (BER או NER), אינדוקציה של חוסר איזון בין הדור וההסרה של נגעים DNA לטובת מוביל לשעבר לעלייה נטו של רמות שלהם ב-DNA שעות נוספות ב-דנ א3 . תוצאות הן הגידול של שיעורי מוטציה DNA, מופחתת ביטוי גנים, ופעילות חלבון מופחת2,4,5,6,7, אפקטים הקשורים היטב ה פיתוח מחלות. מוטציות DNA עשוי להשפיע על פונקציות סלולריות מגוונות, כגון איתות התא, מחזור התא, שלמות הגנום, היציבות טלומר, epigenome, מבנה כרומטין, שחבור RNA, הומאוסטזיס חלבון, מטבוליזם, אפופטוזיס, ובידול התא8 ,9. אסטרטגיות להאט את קצב מוטציה התא ופיתוח מחלות כרוניות (למשל, סרטן, מחלות ניווניות) לעבור דרך הידע של מקורות מוטציה, ביניהם, נגעים DNA והגורמים שלהם.
ROS שנוצר באופן שולי עודף, בשל חשיפה מזהמים, דלקת מתמשכת, פתופסולוגיה המחלה (למשל, סוכרת), וכו ‘, הם גורמים חשובים של נזק ביואואלי, כולל DNA ו נזק לשומנים1. כדוגמה, הידרוקסיל תגובתי מאוד רדיקלי (OH) נוצר מ2O2 הפחתה על ידי מעבר יונים מתכת (פה2 +, Cu+) מחמצן בסיסים dna, moiety סוכר dna ו חומצות שומן רב בלתי רווי ב דיפוזיה מבוקרת מחירים10. בקרב 80 מאופיין בנוקלאואוסים מחמצנים3, הנחקר ביותר הוא 8-אוקסו-7, 8-דיהידרוטסטוסטרון (8-oxogua) או 8-אוקסו-7, 8-dihydro ידרו-2′-deoxyguanosine (8-oxogua, איור 1), נגע כי הוא מסוגל לגרום ל-GT transversions ב תאים ממגלית10,11. הוא נוצר על ידי חמצון אלקטרונית מונו של גואנין, או על ידי רדיקלים הידרוקסיל או סינגרז התקפת חמצן של גואנין ב-DNA1. חומצות שומן רב בלתי רוויות הם מטרות חשובות אחרות של חמצון תגובתי מאוד, כגון •או, אשר ליזום את תהליך של השומנים peroxidation1,12. זה מעניק לחומצות שומן הידרופרתחמוצות שעלולות להתפרק לאלקטרופיפילית אלדדס ואפוקסידז, כגון malondialdehyde, 4-הידרוxy-2-nonenal, 2, 4-דאדיאנאל, 4, 5-אפוקסי-(2ה)-decenal, הקסאנאל, אקרונליין, קרוטונלדהיד, אשר הם מסוגל ליצור מוטאוציציציציציציציציציציציציציציציציציציציציציציציציציציציציציקציציציציציציציציציציציצילציקציקציציציציק האתאי adducts 1,N2-etheno-2′-Deoxyguanosine (1,n2-εdGuo, איור 1) ו-1,n6-etheno-2′-deoxyadenosine (1,n6-εdAdo, איור 1 ) הוצעו כסמנים פוטנציאליים ביוארגיולוגיה של הדלקת14,15.
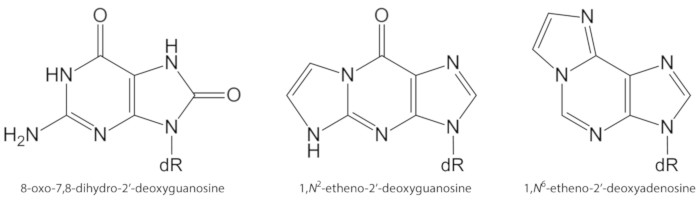
איור 1. מבנים כימיים של נגעים DNA לכמת במחקר הנוכחי. . ד ר 2-דיאוקסיבוז דמות זו השתנתה מתוך אוליביירה ואח ‘.34. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
מחקרים שבוצעו בתחילת שנות השמונים אפשרו זיהוי רגיש של 8-oxodGuo על ידי ביצועים גבוהים כרומטוגרפיה נוזלית מצמידים לזיהוי אלקטרוכימי (כרומטוגרפיה-ECD). הכמת של 8-oxodguo על ידי כדיקות-ECD במספר מערכות ביולוגיות נתון התנאים אוקסיגון הובילו הכרה של 8-oxodguo כמו ביואריקר של oxidatively המושרה נזק DNA1,16. למרות שהוא חזק ומאפשר את הכמת של 8-oxodGuo בטווח הנמוך למול17, כרומטוגרפיה-מדידות ECD להסתמך על הדיוק של זמן השמירה של אנליטה לזיהוי וברזולוציה של כרומטוגרפיה כדי למנוע הפרעות של רכיבים אחרים לדוגמה. כפי שזיהוי אלקטרוכימי דורש שימוש במלח (למשל, אשלגן פוספט, סודיום אצטט) בשלב הנייד, התחזוקה של מצבים אנליטיים נאותים זקוקה לזמן שגרתי של טור וניקוי ציוד.
לחילופין, השימוש של ה-dna בקטריאלי תיקון אנזים מערכת ה-dna גליפסיקלז (fpg) ו, לאחר מכן, האדם 8-oxoguanine גליסיקלז 1 (hOGG1), לאיתור והסרה של 8-oxoגואה מ-dna, יצא כדרך האינדוקציה של dna אלקליות יציב משך אתרים. האתרים האלקליות מומרים למעברי השילוב של דנ א ומאפשרים לכמת את האופיקציה העקיפה הגבוהה ביותר של 8-oxoGua על ידי אלקטרופורזה ביחידה בודדת של ג’ל (“שביט שומר”). את הרגישות הגבוהה ואת ההישג של הניתוחים ללא צורך בחילוץ ה-DNA הסלולר הם היתרונות העיקריים של סוג זה של שיטת. זה נותן את הנמוך ביותר יציב המדינה רמות של 8-oxogua ב-DNA, בדרך כלל 7-10 פעמים נמוך יותר מאשר רמות המתקבלות על ידי שיטות יואנליטיים המבוסס על הבדיקות. עם זאת, היא מדידה עקיפה של 8-oxogua וכמה החסרונות הם חוסר ספציפיות או יעילות לא ידוע של אנזימים תיקון בשימוש1,16,18.
חיסוני הם מערכת אחרים של שיטות המשמשות לזיהוי של 8-האוקגואה1 ו-DNA האקסוציציציציציציציציציציציציצינו1, כגון 1,N6-דאדו ו -1,N2-dguo12. למרות הרגישות, קיצור של השימוש בנוגדנים לאיתור נגעים DNA הוא חוסר ספציפיות בשל פעילות צולבת לרכיבים אחרים של דגימות ביולוגיות, כולל DNA נורמלי בסיסים1,12. כולל דנ א האקסוציטוציציציאלי, לרבות 1,N6-דאדו ו-1,n2-dguo, ניתן גם לזהות ולכמת על ידי רגיש מאוד 32P-postlabeling בחני12. רגישות גבוהה של 32P-התיוג מאפשר את השימוש של כמויות קטנות מאוד של DNA (g., 10 μg) לאיתור של כ 1 קרוב לכל 1010 בסיסים נורמליים19. עם זאת, השימוש בחומרים כימיים, חוסר ספציפיות כימית ודיוק נמוך הם כמה חסרונות19,20.
מגבלה משותפת של השיטות המצוטטות לעיל היא הסלקטיביות הנמוכה או הספציפיות לאיתור המולקולות הרצויות. בתרחיש זה, מצמידים מערכות המדידה לאלקטרו-מגנון מסוג ספקטרומטר המסה (בדיקות-ESI-MS/MS ו-בדיקות-MS3) התפתחו כסטנדרט הזהב לכמת של נוקלאוטידים ששונו בתוך מטריצות ביולוגיות, כגון דנ א, שתן, פלזמה ורוק מיכל בן , מיכל בן 19 , 20. היתרונות של כרומטוגרפיה-ESI-ms/ms הם רגישות (בדרך כלל בטווח fmol נמוך) ואת הספציפיות שסופקו על ידי i) את ההפרדה כרומטוגרפי, ii) תבנית מאפיין וידוע של פיצול מולקולה בתוך המסה תא התנגשות ספקטרומטר, ו iii) את המדידה המדויקת של המסה שנבחרה לחייב את היחס (m/z) במצב ניטור תגובה מרובה1,19. השימוש בתקנים פנימיים המסומנים בisotopically מוסיף את היתרון של תיקונים להפסדים של מולקולה במהלך ההידרוליזה DNA ושלבי העשרת האנליט, כמו גם להבדלים בין הדגימות. זה גם מסייע בזיהוי של שיא כרומואטוגרפי הנכון כאשר יותר משיא אחד קיים1,12,19,20.
מספר שיטות המבוססות על בדיקות – ESI-ms/ms שימשו עבור כימות של 8-oxodguo, 1,N6-דאדו ו 1,n2-dguo ב-DNA מופק דגימות ביולוגיות שונות12,15,20 ,21,22,23,24,25,26,27,28,29 . חלקיקים עדינים (PM2.5) לשאת כימיקלים אורגניים ובלתי אורגניים, כגון פחמימנים ארומטיים רב-מחזורי (pahs), ניטרו-pahs, אלדיודס, ketones, חומצות קרבוקסיליות, quinolines, מתכות, ויונים מסיסים במים, אשר עשויים לגרום לדלקת ו לחץ חמצוני, התנאים שעדיף על התרחשות של נזק ביואואלי ומחלה30,31,32,33. אנו מציגים כאן אימות מאומת-ESI-שיטות ms/ms אשר הוחלו בהצלחה עבור כימות של 8-oxodguo, 1,N6-דאדו ו-1,N2-dguo בריאות, כבד וכליות DNA של עכברים/J להערכת ה ההשפעות של הסביבה PM2.5 חשיפה34.
