Bazı reaktif oksijen türleri (ROS) DNA üsleri ve deoksiriboz Moiety bazı Karbonlar karbon çift bağları okside edebiliyoruz, oksitlenmiş üsler ve DNA iplikçik tatili üreten1. Azot ve oksijen atomları açısından zengin, negatif olarak şarj edilen bir molekül olarak, DNA aynı zamanda nüktenik siteler (azot ve oksijen) ile tepki veren aromatiklerin elektrofilik yerdeğiştirmesi gruplar için bir hedeftir, DNA adduct2denilen ürünler vererek. Yani, DNA adduct ve oksitlenmiş DNA üsleri, elektrofilik olan maddelerin toksisite değerlendirmesi için yararlı biyomarker olan DNA lezyonları örnekleridir, Biyotransformasyon üzerine reaktif elektrofil üretir, ya da oksidatif stres1, 2‘ ye kadar. Değiştirilmiş DNA üsleri DNA ‘dan baz veya nükleotit eksizyon onarımı (BER veya NER) tarafından kaldırılabilse de, DNA lezyonlarının eski yol açmasıyla birlikte DNA lezyonları arasında bir dengesizlik indüksiyonu, daha fazla zaman içinde kendi düzeylerinde net bir artışa neden olur3 . Sonuçlar DNA mutasyonu oranlarının artması, azaltılmış gen ifadesi ve azalmış protein aktivitesi2,4,5,6,7, etkileri ile yakından ilgili olan hastalıkların gelişimi. DNA mutasyonları, hücre sinyalizasyon, hücre döngüsü, genom bütünlüğü, telomer stabilitesi, epigenom, kromatin yapısı, RNA birleştirme, protein homeostasis, metabolizma, apoptoz ve hücre farklılaşma gibi çeşitli hücresel fonksiyonları etkileyebilir8 ,9. Hücre mutasyonu oranlarını ve kronik hastalık gelişimini yavaşlatma stratejileri (örn. kanser, nörodejeneratif hastalıklar) mutasyon kaynaklarının bilgisine, DNA lezyonları ve nedenleri arasında geçmektedir.
Kirletici pozlama, kalıcı inflamasyon, hastalık patofizyolojisi (örn. diyabet) vb. nedeniyle endojenously fazlalıkta oluşturulan Ros, DNA ve lipid hasarı dahil olmak üzere biyomolekül hasarının önemli nedenleridir1. Örnek olarak, H2O2 ‘ den oluşan son derece reaktif hidroksil radikal (Oh) geçiş metal iyonlarına (Fe2 +, cu+) göre difüzyon kontrollü DNA üsleri, DNA şekeri kısım ve çoklu doymamış yağ asitleri oksitlenir oranları10. 80 arasında zaten oksitlenmiş nükvanoüsler karakterize3, en çok çalışan 8-OXO-7, 8-dihidroguanin (8-oxoGua) veya 8-OXO-7, 8-dihydro-2′-deoxyguanosine (8-oxodGuo, Şekil 1), gt transversions içinde yol açan bir lezyon memelinin hücreleri10,11. Bu, guanin mono elektronik oksidasyonu veya DNA1‘ de guanin hidroksil radikal veya atlet oksijen saldırısı ile oluşur. Çoklu doymamış yağ asitleri, lipid peroksidasyonu1,12sürecini başlatan •Oh gibi yüksek reaktif oksidanların diğer önemli hedefleridir. Aromatiklerin elektrofilik yerdeğiştirmesi aldehitler ve epoxyaldehydes için deforme olabilir yağ asidi hidroperoksitler artış sağlar, Malondialdehit, 4-hidroksi-2-nonenal, 2, 4-decadienal, 4, 5-Epoxy-(2E)-decenal, hexenal, acrolein, crotonaldehyde gibi malondialdehit-, Propano-veya etheno adduct1,12,13gibi mutajisik exokiktik DNA adduct ‘ları oluşturabilecektir. Eteno adduct 1,n2-etheno-2′-deoxyguanosine (1,n2-εdguo, Şekil 1) ve 1,n6-etheno-2′-deoxyadenosine (1,n6-εdado, Şekil 1 ) inflamasyonun patofizyolojisinde potansiyel Biyomarkörler olarak önerilen14,15.
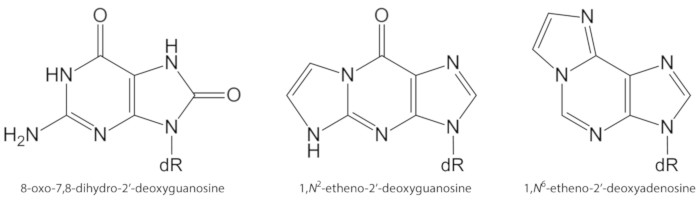
Şekil 1. DNA lezyonları mevcut çalışmada nicelik kimyasal yapıları. dR = 2 ́-deoksirriboz. Bu rakam Oliveira ve al.34güncellenmiştir. Bu figürün daha büyük bir versiyonunu görmek Için lütfen tıklayınız.
1980 ‘ lerin başlarında yürütülen çalışmalar yüksek performanslı sıvı kromatografi elektrokimyasal algılama (HPLC-ECD) ile birleştiğinde 8-oxodGuo hassas tespiti izin verdi. 8-oxodguo tarafından HPLC-ECD tarafından oksitlenme koşullarına maruz kalan çeşitli biyolojik sistemlerde, 8-oxodguo ‘ nın DNA ‘da oksidatif olarak indüklenen baz hasarının bir biyomarker olarak tanınmasına yol açan1,16. Düşük fmol aralığı17‘ de 8-oxodGuo miktarının sağlam olmasına rağmen, HPLC-ECD ölçümleri analitik kimlik tespiti için analit tutma süresi doğruluğunu ve Kromatografi çözünürlüğünü diğer örnek bileşenleri. Elektrokimyasal algılama mobil aşamada tuz kullanımı (örneğin, potasyum fosfat, sodyum asetat) gerektirdiğinden, yeterli analitik koşulların bakımı rutin sütun ve ekipman temizleme süresi gerekir.
Alternatif olarak, bakteriyel DNA onarım enzim formayidopyrimidine DNA glycosylase (FPG) kullanımı ve daha sonra, insan 8-oxoguanine glycosylase 1 (hOGG1), algılama ve DNA ‘dan 8-oxogua kaldırılması için, DNA alkali labil indüksiyon için bir yol olarak ortaya çıktı Site. Alkali labil siteleri DNA Strand molaları dönüştürülür ve alkalin tek hücreli jel elektroforez (“kuyruklu yıldız tahlil”) ile 8-oxogua çok yüksek duyarlı dolaylı kantifikasyon sağlar. Yüksek hassasiyet ve hücresel DNA ekstraksiyon gerek kalmadan analizlerin başarısı bu tür tahlil ana avantajlarıdır. DNA ‘da 8-oxoGua en düşük sabit devlet seviyeleri verir, genellikle 7-10 kez HPLC dayalı biyoanalitik Yöntemler ile elde edilen düzeylerinden daha düşük. Ancak, bu 8-oxogua dolaylı bir ölçüsüdür ve bazı dezavantajları özgüllük eksikliği veya onarım enzimlerin bilinmeyen verimliliği kullanılan1,16,18.
Bağışıklık sistemi, 1,n6-DAdo ve 1,n2-Dguo12gibi 8-oxogua1 ve exokiktik DNA kanalının saptanması için kullanılan diğer yöntemler kümesidir. Duyarlılık rağmen, DNA lezyonlarının tespiti için antikorların kullanımı bir eksiklik biyolojik numunelerin diğer bileşenlere çapraz reaktivite nedeniyle özgüllük eksikliği, normal DNA üsleri dahil,1,12. 1,n6-DAdo ve 1,n2-DGUO dahil olmak üzere exokiklik DNA adduct ‘lar da tespit edilebilir ve son derece hassas 32P-postetiketleme12ile ölçülebilir. 32P-postetiketleme yüksek duyarlılık 1010 normal üsler başına yaklaşık 1 yaklaştırmak tespiti için DNA (örneğin, 10 μg) çok az miktarda kullanımını sağlar19. Ancak, radyo-kimyasallar, kimyasal özgüllük ve düşük doğruluk eksikliği kullanımı bazı dezavantajları19,20.
Yukarıda bahsedilen yöntemlerin paylaşılan sınırlaması, istenilen moleküllerin algılanması için düşük seçicilik veya özgüllük şeklindedir. Bu senaryoda, elektrosprey iyonizasyon tandem kütle spektrometresi (HPLC-ESI-MS/MS ve HPLC-MS3) ile BIRLEŞEN HPLC, DNA, idrar, plazma ve tükürük gibi biyolojik matrislerde değiştirilmiş nükleslerin ölçülmesini için altın standart olarak gelişti. 1 , 19 , 20. HPLC-ESı-MS/MS yöntemlerinin avantajları (genellikle düşük fmol aralığında) duyarlılık ve ı tarafından sağlanan yüksek özgüllük) kromatografik ayrım, II) kütle içinde molekül parçalanma karakteristik ve bilinen deseni Spektrometre çarpışma odası ve III) birden fazla reaksiyon izleme modunda seçilen kütlenin şarj oranı (m/z) doğru ölçümü1,19. İzotopik etiketli iç standartların kullanımı, DNA hidroliz ve analit zenginleştirme adımları sırasında molekül kayıplarına yönelik düzeltmeler ve numuneler arasındaki analit iyonizasyon farklılıkları için bir avantaj katıyor. Aynı zamanda birden fazla zirve mevcut olduğunda doğru kromatografik zirvede teşhis yardımcı olur1,12,19,20.
HPLC-ESI-MS/MS bazlı çeşitli yöntemler, farklı biyolojik örneklerden çıkarılan DNA ‘da 8-oxodguo, 1,n6-dAdo ve 1,n2-dguo miktarının ölçülmesini için kullanılmıştır12,15,20 ,21,22,23,24,25,26,27,28,29 . İnce parçacıklar (PM2,5) organik ve inorganik kimyasallar taşır, polisiklik aromatik hidrokarbonlar (PAH) gibi, Nitro-PAHs, aldehyler, ketonlar, karboksilik asitler, kinolinler, metaller, ve su çözünür iyonlarının, hangi inflamasyon neden olabilir ve oksidatif stres, biyomolekül hasar ve hastalığın oluşumu lehine koşullar30,31,32,33. Burada onaylanan HPLC-ESı-MS/MS yöntemleri, 8-oxodGuo, 1,n6-DAdo ve 1,n2-DGUO in Lung, karaciğer ve böbrek DNA ‘sı/J farelerinin ölçülmesi için başarıyla uygulanmıştır. ortam PM2,5 pozlama34etkileri.
