Nogle reaktive oxygenarter (ROS) er i stand til at oxideres Carbon dobbeltbindinger af DNA-baser og nogle kulstofatomer i Deoxyribose-delen, genererer oxiderede baser og DNA-streng pauser1. Som et negativt ladet molekyle, der er rigt på nitrogen og oxygen atomer, er DNA også et mål for elektro File grupper, der kovalent reagerer med de nukleofile steder (nitrogen og ilt), hvilket giver produkter, der kaldes DNA-addukter2. Så, DNA addukter og oxiderede DNA-baser er eksempler på DNA-læsioner, der er nyttige biomarkører for toksicitets vurdering af stoffer, der er elektro File, generere reaktive elektro philes ved biotransformation, eller inducere oxidativ stress1, 2. i. Selv om de modificerede DNA baser kan fjernes fra DNA ved base eller nukleotid excision reparation (BER eller NER), induktion af en ubalance mellem generering og fjernelse af DNA-læsioner til fordel for førstnævnte fører til en nettostigning i deres niveauer i DNA overarbejde3 . Resultater er stigningen i DNA-Mutations rater, reduceret genekspression og formindsket protein aktivitet2,4,5,6,7, virkninger, der er tæt forbundet med udvikling af sygdomme. DNA-mutationer kan påvirke forskellige cellulære funktioner, såsom celle signalering, celle cyklus, genomintegritet, telomere stabilitet, epigenom, kromatin struktur, RNA splejsning, protein homøostase, metabolisme, apoptose og Celledifferentiering8 ,9. Strategier til at bremse celle Mutations rater og kronisk sygdomsudvikling (f. eks. kræft, neurodegenerative sygdomme) passerer gennem viden om Mutations kilderne, heriblandt DNA-læsioner og deres årsager.
ROS genereret endogent i overskud, på grund af forurening eksponering, vedvarende inflammation, sygdom Patofysiologi (f. eks diabetes), etc., er vigtige årsager til biomolekyle skade, herunder DNA og lipid skader1. Som et eksempel er den meget reaktive hydroxyl radikal (OH) dannet af H2O2 reduktion af Transition metalioner (fe2 +, cu+) oxiderer DNA baserne, DNA-sukker delen og flerumættede fedtsyrer ved diffusions kontrolleret priser10. Blandt de 80 allerede karakteriseret oxiderede nukleobaser3, den mest undersøgt ene er 8-oxo-7, 8-dihydroguanin (8-oxogua) eller 8-oxo-7, 8-dihydro-2′-deoxyguanosin (8-oxodGuo, figur 1), en læsion, der er i stand til at inducere gt Trans versioner i pattedyrsceller10,11. Det dannes ved mono elektronisk oxidation af guanin eller ved hydroxyl radikal eller singlet oxygen angreb af guanin i DNA1. Flerumættede fedtsyrer er andre vigtige mål for meget reaktive oxidanter, såsom •Oh, som indleder processen med lipid peroxidering1,12. Det giver anledning til fedtsyre hydroperoxider, der kan nedbrydes til elektro File aldehyder og epoxyaldehyder, såsom malondialdehyd, 4-hydroxy-2-nonenal, 2,4-decadienal, 4,5-epoxy-(2E)-decenal, hexenal, acrolein, crotonaldehyd, som er kunne danne mutagene exocycliske DNA-addukter, såsom malondialdehyd-, Propano-eller etheno-addukter1,12,13. Etheno adkanalerne 1,n2-etheno-2′-deoxyguanosin (1,n2-εdguo, figur 1) og 1,n6-etheno-2′-deoxyadenosin (1,n6-εdado, figur 1 ) er blevet foreslået som potentielle biomarkører i Patofysiologi af inflammation14,15.
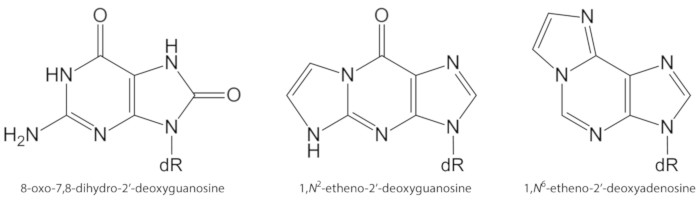
Figur 1. Kemiske strukturer af de DNA-læsioner, der er kvantificeret i nærværende undersøgelse. dR = 2 ́-Deoxyribose. Dette tal er blevet ændret fra Oliveira et al.34. Klik her for at se en større version af dette tal.
Undersøgelser foretaget i begyndelsen af 1980 ‘ erne tillod følsom påvisning af 8-oxodGuo ved højtydende væskekromatografi koblet til elektrokemisk detektion (HPLC-ECD). Kvantificering af 8-oxodguo ved HPLC-ECD i flere biologiske systemer udsat for oxiderende betingelser førte til anerkendelse af 8-oxodguo som en biomarkør af oxidativt induceret base skade i DNA1,16. Selv om den er robust og giver mulighed for kvantificering af 8-oxodGuo i lavfmol-området17, afhænger HPLC-ECD-målingerne af nøjagtigheden af analysand-retentionstiden for analyt-identifikation og af kromatografi opløsningen for at undgå interferenser med andre prøve bestanddele. Da elektrokemisk detektion kræver brug af salt (f. eks. kaliumphosphat, natriumacetat) i den mobile fase, skal vedligeholdelse af passende analytiske betingelser rutine kolonne og rengøringstiden for udstyr.
Alternativt, brugen af bakteriel DNA reparation enzym formamidopyrimidine DNA glycosylase (FPG) og bagefter, humant 8-oxoguanine glycosylase 1 (hOGG1), til påvisning og fjernelse af 8-oxoGua fra DNA, opstod som en måde for induktion af DNA alkali labile Websteder. De alkaliske labile steder omdannes til DNA-streng pauser og tillader en meget høj følsom indirekte kvantificering af 8-oxoGua ved alkalisk enkelt celle gel elektroforese (“Comet assay”). Den høje følsomhed og udførelsen af analyserne uden behov for cellulær DNA ekstraktion er de vigtigste fordele ved denne type analyse. Det giver de laveste Steady-State niveauer af 8-oxoGua i DNA, typisk 7-10 gange lavere end niveauerne opnået ved bioanalytiske metoder baseret på HPLC. Men det er en indirekte måling af 8-oxogua og nogle ulemper er manglen på specificitet eller den ukendte effektivitet af reparation enzymer brugt1,16,18.
Immunoassays er andre sæt metoder, der anvendes til påvisning af 8-oxoGua1 og exocyclic DNA addukter, såsom 1,n6-Dado og 1,n2-dguo12. På trods af følsomheden er en mangel ved brugen af antistoffer til påvisning af DNA-læsioner manglen på specificitet på grund af kryds reaktivitet til andre bestanddele af biologiske prøver, herunder de normale DNA baser1,12. De exocycliske DNA-addukter, herunder 1, n6-Dado og 1, n2-dguo, kan også påvises og kvantificeres af meget følsomme 32P-post mærknings-analyser12. Den høje følsomhed af 32P-post mærkning tillader brug af meget små mængder af DNA (f. eks 10 μg) til påvisning af omkring 1 adduct per 1010 normale baser19. Men brugen af radio-kemikalier, manglende kemisk specificitet og lav nøjagtighed er nogle ulemper19,20.
En fælles begrænsning af de ovenfor citerede metoder er den lave selektivitet eller specificitet for påvisning af de ønskede molekyler. I dette scenario er HPLC koblet til elektro spray ionisering tandem massespektrometri (HPLC-ESI-MS/MS og HPLC-MS3) udviklet som guld standard for kvantificering af modificerede Nukleosider i biologiske matricer, såsom DNA, urin, plasma og spyt 1 af , 19 ud af , 20. fordele ved HPLC-ESI-MS/MS metoder er følsomheden (typisk i den lave fmol Range) og den høje specificitet, som i) kromatografi separation, II) det karakteristiske og kendte mønster af molekyle fragmentering inde i massen spektrometer kollisions kammer og III) nøjagtig måling af den valgte masse til opladnings forhold (m/z) i flere reaktions overvågnings modus1,19. Brugen af isotopisk mærkede interne standarder tilføjer fordelen ved korrektioner for Molekylær tab under DNA-hydrolyse-og analyt-berignings trinnene samt for forskelle i analytioniseringen mellem prøverne. Det bidrager også til identifikationen af den korrekte kromatografi, når der er mere end én top til stede1,12,19,20.
Flere metoder baseret på HPLC-ESI-MS/MS er blevet anvendt til kvantificering af 8-oxodguo, 1, n6-Dado og 1,N2-dguo i dna ekstraheret fra forskellige biologiske prøver12,15,20 ,21,22,23,24,25,26,27,28,29 . Fine partikler (PM2,5) transporterer organiske og uorganiske kemikalier, såsom polycykliske aromatiske kulbrinter (PAH), nitro-PAH’er, aldehyder, ketoner, carboxylsyrer, quinoliner, metaller og vandopløselige ioner, som kan inducere betændelse og oxidativ stress, betingelser, der begunstiger forekomsten af biomolekyle skader og sygdom30,31,32,33. Vi præsenterer her validerede HPLC-ESI-MS/MS-metoder, der med succes blev anvendt til kvantificering af 8-oxodGuo, 1,n6-Dado og 1,N2-dguo i LUNGE-, lever-og nyre-DNA fra A/J-mus til vurdering af virkninger af omgivelsernes PM2,5 -eksponering34.
