Vissa reaktiva syreradikaler (ROS) kan oxidera kol dubbel bindningar av DNA-baser och vissa kol atomer i deoxyribose del, generera oxiderade baser och DNA strand raster1. Som en negativt laddad molekyl rik på kväve-och syre atomer är DNA också ett mål för elektrofila grupper som kovalent reagerar med de nukleofila platserna (kväve och syre), vilket ger produkter som kallas DNA-addukter2. Så, DNA addukter och oxiderade DNA baser är exempel på DNA-lesioner som är användbara bio markörer för toxicitet bedömning av ämnen som är elektrofila, generera reaktiva elektrophiles på bio Transformation, eller inducera oxidativ stress1, och 2. Även om de modifierade DNA-baser kan avlägsnas från DNA genom bas eller nukleotid excision reparation (BER eller NER), induktion av en obalans mellan generering och avlägsnande av DNA-lesioner till förmån för de förstnämnda leder till en nettoökning av deras nivåer i DNA övertid3 . Resultat är ökningen av DNA-mutations frekvenser, minskat gen uttryck och minskad protein aktivitet2,4,5,6,7, effekter som är nära relaterade till utveckling av sjukdomar. DNA-mutationer kan påverka olika cellulära funktioner, såsom cellsignalering, cell cykel, genom integritet, telomer stabilitet, epigenomet, kromatin struktur, RNA splitsning, protein homeostas, metabolism, apoptos, och cell differentiering8 ,9. Strategier för att bromsa cell mutations frekvenser och kronisk sjukdoms utveckling (t. ex. cancer, neurodegenerativa sjukdomar) passerar genom kunskapen om mutationskällorna, bland dem, DNA-lesioner och deras orsaker.
ROS genereras endogent i överskott, på grund av förorening exponering, ihållande inflammation, sjukdomspatofysiologi (t. ex., diabetes), etc., är viktiga orsaker till bio molekyl ära skador, inklusive DNA och lipid skador1. Som ett exempel, den mycket reaktiva hydroxylradikal (OH) bildas från H2O2 reduktion av över gång metall joner (Fe2 +, cu+) oxiderar DNA baser, DNA-socker delen och fleromättade fett syror vid diffusions-kontrollerad och räntor10. Bland de 80 redan karakteriserade oxiderade nukleobaser3, den mest studerade en är 8-oxo-7, 8-dihydroguanin (8-oxoGua) eller 8-oxo-7, 8-dihydro-2 ‘-deoxyguanosin (8-oxodGuo, figur 1), en lesion som kan inducera gt transversioner i däggdjurs celler10,11. Det bildas genom mono elektronisk oxidation av guanin, eller genom hydroxylgrupp radikal eller singlet syre attack av guanin i DNA1. Fleromättade fett syror är andra viktiga mål för mycket reaktiva oxidanter, såsom •Oh, som initierar processen för lipidperoxidation1,12. Det ger upphov till fett syra Hydro per oxider som kan sönderdelas till elektrofila aldehyder och epoxyaldehyder, såsom malondialdehyd, 4-hydroxi-2-nonenal, 2, 4-decadienal, 4,5-epoxi-(2E)-decenal, hexenal, acrolein, crotonaldehyd, som kunna bilda mutagena exocykliska DNA-addukter, såsom malondialdehyd-, Propano-, eller etheno addukter1,12,13. Etheno addukter 1,n2-etheno-2 ‘-deoxyguanosin (1,n2-εdguo, figur 1) och 1,n6-etheno-2 ‘-Deoxyadenosin (1,n6-εdado, figur 1 ) har föreslagits som potentiella bio markörer i patofysiologin av inflammation14,15.
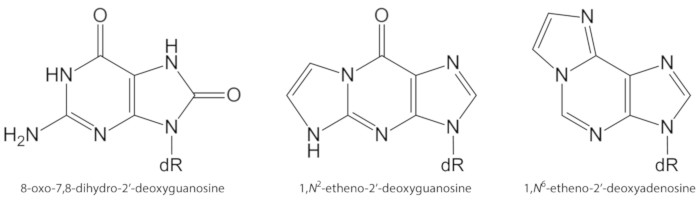
I figur 1. Kemiska strukturerna hos de DNA-lesioner som kvantifierats i den här studien. dR = 2 ́-deoxyribose. Denna siffra har ändrats från Oliveira et al.34. Vänligen klicka här för att se en större version av denna siffra.
Studier som genomfördes i början av 1980-talet tillät den känsliga upptäckten av 8-oxodGuo av högpresterande vätskekromatografi kopplad till elektrokemisk detektion (HPLC-ECD). Kvantifiering av 8-oxodguo av HPLC-ECD i flera biologiska system utsätts för oxiderande förhållanden ledde till erkännandet av 8-oxodguo som en bio markör av oxidativt inducerad bas skador i DNA1,16. Även om det är robust och gör det möjligt att kvantifiera 8-oxodGuo i det låga fmol-området17, förlitar sig HPLC-ECD-mätningar på noggrannheten hos analytretentions tiden för analytidentifiering och på kromatografiupplösningen för att undvika interferenser av andra prov bestånds delar. Eftersom den elektrokemiska upptäckten kräver användning av salt (t. ex. kalium fosfat, natriumacetat) i den mobila fasen, behöver underhållet av lämpliga analytiska tillstånd rutinmässig kolonn och utrustning rengörings tid.
Alternativt, användning av bakterien DNA reparation enzym formamidopyrimidine DNA glykosylase (FPG) och, efteråt, mänskliga 8-oxoguanin glykosylase 1 (hOGG1), för detektion och avlägsnande av 8-oxogua från DNA, framkom som ett sätt för induktion av DNA alkali labila Platser. De alkalilabila platserna konverteras till DNA-strandbryt och tillåter mycket hög känslig indirekt kvantifiering av 8-oxogua genom alkalisk Single cell gel elektrofores (“kometanalys”). Den höga känsligheten och utförandet av analyserna utan behov av cellulära DNA-extraktion är de främsta fördelarna med denna typ av analys. Det ger den lägsta Steady-State nivåer av 8-oxoGua i DNA, typiskt 7-10 gånger lägre än de nivåer som erhålls genom bio analytiska metoder baserade på HPLC. Det är dock en indirekt mätning av 8-oxogua och vissa nack delar är bristen på specificitet eller den okända effektiviteten i reparera enzymer som används1,16,18.
Immunoassay är andra uppsättning metoder som används för detektion av 8-oxoGua1 och EXOCYKLISKA DNA addukter, såsom 1,n6-Dado och 1,n2-dguo12. Trots känsligheten, en brist på användning av anti kroppar för detektion av DNA-lesioner är bristen på specificitet på grund av korsreaktivitet till andra komponenter i biologiska prover, inklusive de normala DNA-baser1,12. Den exocykliska DNA addukter, inklusive 1,n6-Dado och 1,n2-dguo, kan också upptäckas och kvantifieras av mycket känsliga 32P-postmärkning analyser12. Den höga känsligheten hos 32P-postmärkning gör det möjligt att använda mycket små mängder DNA (t. ex. 10 μg) för detektion av ca 1 ankring per 1010 normal baser19. Emellertid, bruket av Radios ände-kemikalieer, brist av kemisk specificitet och låg exakthet är några missgynnar19,20.
En delad begränsning av de metoder som nämns ovan är den låga selektivitet eller specificitet för detektion av de önskade molekylerna. I det här scenariot utvecklades HPLC kopplat till elektrospray jonisering tandem masspektrometri (HPLC-ESI-MS/MS och HPLC-MS3) som guld standard för kvantifiering av modifierade nukleosidor i biologiska matriser, såsom DNA, urin, plasma och saliv 1 den första , den 19 , 20. fördelar med HPLC-ESI-MS/MS-metoder är känsligheten (vanligt vis i det låga fmol-intervallet) och den höga specificitet som i) den kromatografiska separationen, II) det karakteristiska och kända mönstret av molekylfragmentering i massan spektrometer kollision kammare, och III) exakt mätning av den valda massan att ladda förhållandet (m/z) i flera reaktions övervakning läge1,19. Användningen av isotopiskt märkta interna standarder ger fördelen av korrigeringar för molekyl förluster under DNA hydrolys och analyten anriknings steg, samt för skillnader i analytjonisering mellan proverna. Det hjälper också i identifieringen av den korrekta kromatografiska toppen när mer än en topp är närvarande1,12,19,20.
Flera metoder baserade på HPLC-ESI-MS/MS har använts för kvantifiering av 8-oxodguo, 1,n6-Dado och 1,n2-dguo i DNA som extraherats från olika biologiska prover12,15,20 ,21,22,23,24,25,26,27,28,29 . Fina partiklar (PM2,5) bär organiska och oorganiska kemikalier, såsom polycykliska aromatiska kolväten (PAH), Nitro-PAH, aldehyder, ketoner, karboxylsyror, kinoliner, metaller och vattenlösliga joner, som kan framkalla inflammation och oxidativ stress, förhållanden som gynnar förekomsten av bio molekyl skador och sjukdomar30,31,32,33. Vi presenterar här validerade HPLC-ESI-MS/MS-metoder som framgångs rikt tillämpades för kvantifiering av 8-oxodGuo, 1,n6-Dado och 1,n2-dguo i lung-, lever-och njure DNA från A/J-möss för bedömning av effekter av ambient PM2,5 exponering34.
