Een groot aantal modellen zijn beschikbaar om de progressie van kanker te bestuderen, elk van hen is uniek en vertegenwoordigt een subtype van deze complexe ziekte. Elk model biedt unieke en waardevolle inzichten in de biologie van kanker en heeft de middelen verbeterd om de werkelijke ziektetoestand na te bootsen. Gevestigde cellijnen die als monolaag zijn gegroeid, hebben waardevolle inzichten opgeleverd in vitale processen in vitro, zoals proliferatie, invasiviteit, migratie en apoptose1. Hoewel tweedimensionale (2D) celcultuur het traditionele hulpmiddel is geweest om de reactie van zoogdiercellen op verschillende omgevingsverstoringen te onderzoeken, lijkt extrapolatie van deze bevindingen om reacties op weefselniveau te voorspellen niet overtuigend genoeg. De belangrijkste beperking van de 2D-culturen is dat de gecreëerde micro-omgeving grotendeels verschilt van die van het borstweefsel zelf2. 2D-cultuur mist de interactie van de cellen met de extracellulaire matrix, die van vitaal belang is voor de groei van elk weefsel. Ook belemmeren trekkrachten die de cel in monolaagculturen ervaart de polariteit van deze cellen, waardoor de celsignalering en het gedrag veranderen 3,4,5. Driedimensionale (3D) kweeksystemen hebben een nieuwe weg geopend op het gebied van kankeronderzoek met hun vermogen om de in vivo omstandigheden in vitro na te bootsen. Veel cruciale micro-omgevingssignalen die verloren gaan in 2D-celcultuur kunnen worden hersteld met behulp van 3D-culturen van lamininerijke extracellulaire matrix (lrECM)6.
Verschillende studies hebben het belang van de tumor micro-omgeving in carcinogenese geïdentificeerd 7,8. Ontstekingsgerelateerde factoren zijn een belangrijk onderdeel van de micro-omgeving. Platelet Activating Factor (PAF) is een fosfolipidemediator die wordt uitgescheiden door verschillende immuuncellen en die meerdere immuunresponsen bemiddelt 9,10. Hoge niveaus van PAF worden uitgescheiden door verschillende borstkankercellijnen en worden geassocieerd met verhoogde proliferatie11. Studies van ons laboratorium hebben aangetoond dat de langdurige aanwezigheid van PAF in acinaire culturen leidt tot de transformatie van borstepitheelcellen12. PAF activeert de PAF-receptor (PAFR) en activeert de PI3K/Akt-signaleringsas13. PAFR wordt ook geassocieerd met EMT, invasie en metastase14.
Het huidige protocol demonstreert een modelsysteem om PAF-geïnduceerde transformatie te bestuderen, met behulp van 3D-culturen van borstepitheelcellen, zoals eerder is beschreven door Chakravarty et al.12. De borstepitheelcellen die op de extracellulaire matrix (3D-culturen) zijn gegroeid, hebben de neiging om gepolariseerde groei-arresteerde sferoïden te vormen. Deze worden acini genoemd en lijken sterk op de acini van borstweefsel, de kleinste functionele eenheid van de borstklier, in vivo15. Deze sferoïden (figuur 1A,B) bestaan uit een monolaag van dicht opeengepakte gepolariseerde epitheelcellen rond een hol lumen en bevestigd aan het keldermembraan (figuur 1C). Dit proces van morfogenese is goed beschreven in literatuur16. Wanneer ze op lrECM worden gezaaid, ondergaan de cellen deling en differentiatie om een cluster van cellen te vormen, die vervolgens vanaf dag 4 polariseren. Op dag 8 bestaan de acini uit een groep gepolariseerde cellen die in direct contact staan met de extracellulaire matrix en een cluster van niet-gepolariseerde cellen ingesloten in de buitenste gepolariseerde cellen, zonder contact met de matrix. Van deze niet-gepolariseerde cellen is bekend dat ze op dag 12 van de cultuur apoptose ondergaan en een hol lumen vormen. Op dag 16 worden groei-arresteerde structuren gevormd16.
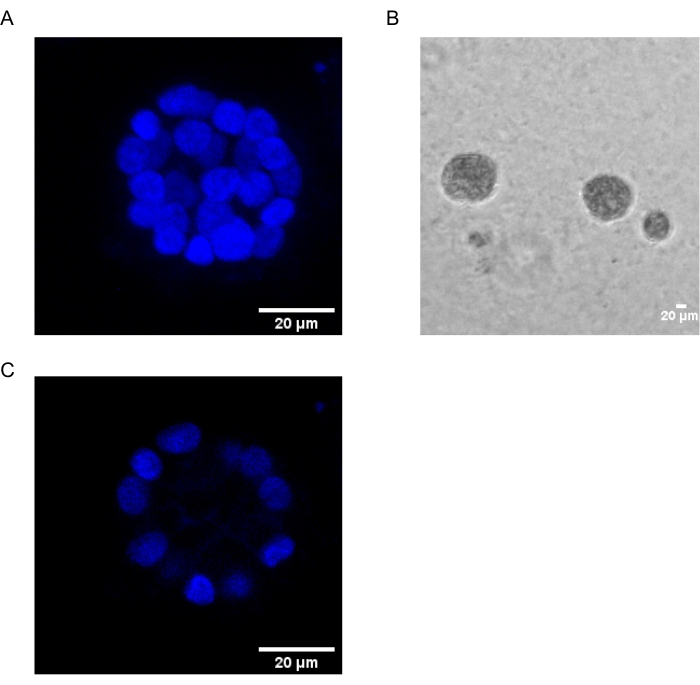
Figuur 1: Kernen van cellen in acini gekleurd met een kernvlek. (A) 3D-constructie van de acini. (B) Fasecontrastbeeld van MCF10A acini gekweekt op Matrigel gedurende 20 dagen. (C) Het middelste gedeelte toont de aanwezigheid van een hol lumen. Schaalbalk = 20 μm. Klik hier om een grotere versie van dit cijfer te bekijken.
In tegenstelling tot 2D-culturen helpen acinarculturen bij het onderscheiden van normale en getransformeerde cellen door middel van schijnbare morfologische veranderingen. Niet-getransformeerde borstepitheelcellen vormen acini met een hol lumen, dat de normale menselijke borst acini nabootst. Deze sferoïden vertonen bij transformatie een verstoorde morfologie die wordt gekenmerkt door een groot verlies van polariteit (een van de kenmerken van kanker), afwezigheid van een lumen of verstoring van het holle lumen (als gevolg van ontwijking van apoptose) die kan worden geïnduceerd als gevolg van deregulatie van verschillende genen 17,18,19,20 . Deze transformaties kunnen worden bestudeerd met behulp van veelgebruikte technieken zoals immunofluorescentie. Het 3D-celkweekmodel kan dus functioneren als een eenvoudige methode om het proces van borstacinaire morfogenese en carcinogenese van de borst te onderzoeken. Het opzetten van een 3D-kweeksysteem om het effect van een fosfolipidemediator, PAF, te begrijpen, zal helpen bij preklinische geneesmiddelenscreening met hoge doorvoer.
Dit werk heeft het 3D ‘on top’ cultuurprotocol16,21 aangepast voor het bestuderen van transformatie geïnduceerd door PAF22. De fenotypische veranderingen veroorzaakt door blootstelling van de acini aan de fosfolipide mediator werden bestudeerd met behulp van immunofluorescentie. Verschillende polariteit en epitheliale naar mesenchymale overgang (EMT) markers12,16 werden gebruikt in de studie. Tabel 1 vermeldt hun normale lokalisatie en hun verwachte fenotype bij transformatie.
| Antilichamen | Merken | Normale lokalisatie | Getransformeerd fenotype |
| α6-Integrin | Basolateraal | Basaal met zwakke laterale vlek | Sterke laterale / apicale vlek |
| β-Catenin | Cel-cel junctie | Basolateraal | Abnormale / nucleaire of cytoplasmatische lokalisatie |
| Vimentin | Emt | Afwezige / zwakke aanwezigheid | Up-regulering |
Tabel 1: Markers gebruikt in het onderzoek. Verschillende markers gebruikt met hun lokalisatie in de aan- en afwezigheid van PAF-behandeling.
Deze methode kan het best worden gebruikt om plausibele geneesmiddelen en doelgenen voor verschillende subtypen borstkanker te bestuderen / screenen. Dit kan een geneesmiddelresponsgegevens opleveren die dichter bij het in vivo scenario liggen, wat helpt bij een snellere en betrouwbaardere ontwikkeling van geneesmiddelen. Dit systeem kan ook worden gebruikt om de moleculaire signalering te bestuderen die verband houdt met medicijnrespons en medicijnresistentie.
