Uma miríade de modelos estão disponíveis para estudar a progressão do câncer, sendo cada um deles único e representando um subtipo dessa doença complexa. Cada modelo fornece insights únicos e valiosos sobre a biologia do câncer e melhorou os meios para imitar a condição real da doença. Linhas celulares estabelecidas cultivadas como monocamadas forneceram insights valiosos sobre processos vitais in vitro, como proliferação, invasividade, migração e apoptose1. Embora a cultura celular bidimensional (2D) tenha sido a ferramenta tradicional para investigar a resposta das células mamíferas a várias perturbações ambientais, a extrapolação desses achados para prever respostas em nível tecidual não parece suficientemente convincente. A maior limitação das culturas 2D é que o microambiente criado difere em grande parte do do próprio tecido mamário2. A cultura 2D carece da interação das células com a matriz extracelular, que é vital para o crescimento de qualquer tecido. Além disso, as forças de tração experimentadas pela célula nas culturas de monocamadas dificultam a polaridade dessas células, alterando assim a sinalização celular e o comportamento 3,4,5. Sistemas de cultura tridimensionais (3D) abriram uma nova avenida no campo da pesquisa sobre câncer com sua capacidade de imitar as condições in vivo in vitro. Muitas pistas microambientais cruciais que são perdidas na cultura celular 2D poderiam ser restabelecidas usando culturas 3D de matriz extracelular rica em laminino (lrECM)6.
Vários estudos identificaram a importância do microambiente tumoral na carcinogênese 7,8. Fatores associados à inflamação são uma parte importante do microambiente. O Fator de Ativação de Plaquetas (PAF) é um mediador fosfolipídide secretado por várias células imunes que media múltiplas respostas imunes 9,10. Altos níveis de PAF são secretados por diferentes linhas de células cancerígenas de mama e estão associados à maior proliferação11. Estudos do nosso laboratório mostraram que a presença prolongada de PAF em culturas acinar leva à transformação das células epiteliaismamárias 12. O PAF ativa o receptor PAF (PAFR), ativando o eixo de sinalização PI3K/Akt13. O PAFR também está associado ao EMT, invasão e metástase14.
O presente protocolo demonstra um sistema modelo para estudar a transformação induzida pelo PAF, utilizando culturas 3D de células epiteliais mamárias, como foi descrito anteriormente por Chakravarty et al.12. As células epiteliais mamárias cultivadas na matriz extracelular (culturas 3D) tendem a formar esferoides polarizados presos pelo crescimento. Estes são chamados de acini e se assemelham muito ao acini do tecido mamário, a menor unidade funcional da glândula mamária, in vivo15. Estes esferoides (Figura 1A,B) consistem em uma monocamada de células epiteliais polarizadas estreitamente embaladas em torno de um lúmen oco e anexadas à membrana do porão (Figura 1C). Este processo de morfogênese foi bem descrito na literatura16. Quando semeadas no LrECM, as células passam por divisão e diferenciação para formar um aglomerado de células, que então polarizam a partir do dia 4. Até o dia 8, o acini consiste em um grupo de células polarizadas que estão em contato direto com a matriz extracelular e um aglomerado de células não polarizadas dentro das células externas polarizadas, sem contato com a matriz. Essas células não polidas são conhecidas por serem submetidas à apoptose até o dia 12 da cultura, formando um lúmen oco. Até o dia 16, as estruturas presas pelo crescimento são formadasem 16.
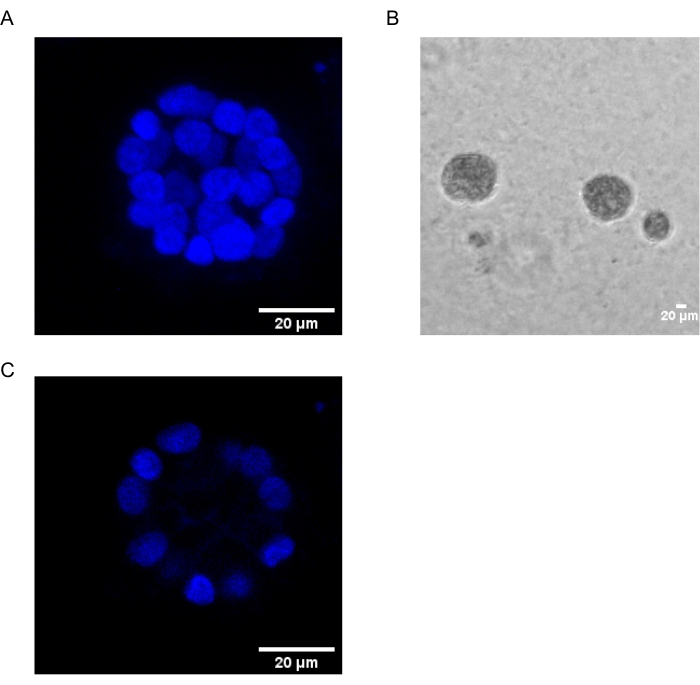
Figura 1: Núcleos de células em acini manchadas com uma mancha nuclear. (A) construção 3D do acini. (B) Imagem de contraste de fase do acini MCF10A cultivada em Matrigel por 20 dias. (C) A seção mais central mostra a presença de um lúmen oco. Barra de escala = 20 μm. Clique aqui para ver uma versão maior desta figura.
Ao contrário das culturas 2D, as culturas acinar ajudam a distinguir células normais e transformadas através de mudanças aparentes de morfologia. Células epiteliais de mama não transformadas formam acini com um lúmen oco, imitando o acini mamário humano normal. Esses esferoides, após a transformação, mostram uma morfologia interrompida caracterizada por uma grande perda de polaridade (uma das marcas do câncer), ausência de lúmen, ou interrupção do lúmen oco (devido à evasão da apoptose) que pode ser induzida devido à desregulamentação de vários genes 17,18,19,20 . Essas transformações podem ser estudadas utilizando técnicas comumente utilizadas, como a imunofluorescência. Assim, o modelo de cultura celular 3D pode funcionar como um método simples para investigar o processo de morfogênese acinar mama e carcinogênese mamária. A criação de um sistema de cultura 3D para entender o efeito de um mediador fosfolipídico, PAF, ajudará na triagem de medicamentos pré-clínicos de alto rendimento.
Este trabalho adaptou o protocolo de cultura 3D ‘no topo’16,21 para estudar a transformação induzida pelo PAF22. As alterações fenotípicas induzidas pela exposição do acini ao mediador fosfolipídico foram estudadas por meio da imunofluorescência. Vários marcadores de polaridade e epitelial para transição mesenquimal (EMT)foram utilizados no estudo. A Tabela 1 menciona sua localização normal e seu fenótipo esperado após a transformação.
| Anticorpos | Marcas | Localização normal | Fenótipo transformado |
| α6-Integrin | Basolateral | Basal com mancha lateral fraca | Mancha lateral forte / Apical |
| β-Catenin | Junção celular | Basolateral | Localização anormal / nuclear ou citoplasmática |
| Vimentina | Emt | Presença ausente/fraca | Up-regulation |
Tabela 1: Marcadores utilizados no estudo. Diferentes marcadores utilizados com sua localização na presença e ausência de tratamento paf.
Este método pode ser melhor utilizado para estudar/testar drogas plausíveis e genes-alvo para vários subtipos de câncer de mama. Isso pode fornecer um dado de resposta a medicamentos mais próximo do cenário in vivo , auxiliando no desenvolvimento mais rápido e confiável de medicamentos. Além disso, este sistema pode ser usado para estudar a sinalização molecular associada à resposta a drogas e resistência a drogas.
