Una miríada de modelos están disponibles para estudiar la progresión del cáncer, cada uno de ellos es único y representa un subtipo de esta compleja enfermedad. Cada modelo proporciona información única y valiosa sobre la biología del cáncer y ha mejorado los medios para imitar la condición real de la enfermedad. Las líneas celulares establecidas cultivadas como una monocapa han proporcionado información valiosa sobre procesos vitales in vitro, como la proliferación, la invasividad, la migración y la apoptosis1. Aunque el cultivo celular bidimensional (2D) ha sido la herramienta tradicional para investigar la respuesta de las células de mamíferos a varias perturbaciones ambientales, la extrapolación de estos hallazgos para predecir las respuestas a nivel tisular no parece lo suficientemente convincente. La principal limitación de los cultivos 2D es que el microambiente creado difiere en gran medida del tejido mamario en sí2. El cultivo 2D carece de la interacción de las células con la matriz extracelular, que es vital para el crecimiento de cualquier tejido. Además, las fuerzas de tracción experimentadas por la célula en cultivos monocapa dificultan la polaridad de esas células, alterando así la señalización celular y el comportamiento 3,4,5. Los sistemas de cultivo tridimensionales (3D) han abierto una nueva vía en el campo de la investigación del cáncer con su capacidad para imitar las condiciones in vivo in vitro. Muchas señales microambientales cruciales que se pierden en el cultivo celular 2D podrían restablecerse utilizando cultivos 3D de matriz extracelular rica en laminina (lrECM)6.
Diversos estudios han identificado la importancia del microambiente tumoral en la carcinogénesis 7,8. Los factores asociados a la inflamación son una parte importante del microambiente. El factor activador plaquetario (PAF) es un mediador fosfolípido secretado por varias células inmunes que media múltiples respuestas inmunes 9,10. Los altos niveles de PAF son secretados por diferentes líneas celulares de cáncer de mama y están asociados con una mayor proliferación11. Estudios de nuestro laboratorio han demostrado que la presencia prolongada de PAF en cultivos acinares conduce a la transformación de las células epiteliales de mama12. PAF activa el receptor PAF (PAFR), activando el eje de señalización PI3K/Akt13. También se informa que el FAP está asociado con EMT, invasión y metástasis14.
El presente protocolo demuestra un sistema modelo para estudiar la transformación inducida por PAF, utilizando cultivos 3D de células epiteliales de mama, como ha sido descrito previamente por Chakravarty et al.12. Las células epiteliales mamarias cultivadas en la matriz extracelular (cultivos 3D) tienden a formar esferoides polarizados con crecimiento detenido. Estos son llamados acinos y se asemejan mucho a los acinos del tejido mamario, la unidad funcional más pequeña de la glándula mamaria, in vivo15. Estos esferoides (Figura 1A, B) consisten en una monocapa de células epiteliales polarizadas estrechamente empaquetadas que rodean una luz hueca y están unidas a la membrana basal (Figura 1C). Este proceso de morfogénesis ha sido bien descrito en la literatura16. Cuando se siembran en lrECM, las células se dividen y diferencian para formar un grupo de células, que luego se polarizan desde el día 4 en adelante. Para el día 8, los acinos consisten en un grupo de células polarizadas que están en contacto directo con la matriz extracelular y un grupo de células no polarizadas encerradas dentro de las células polarizadas externas, sin contacto con la matriz. Se sabe que estas células no polarizadas sufren apoptosis en el día 12 de cultivo, formando una luz hueca. Para el día 16, se forman estructuras con crecimiento detenido16.
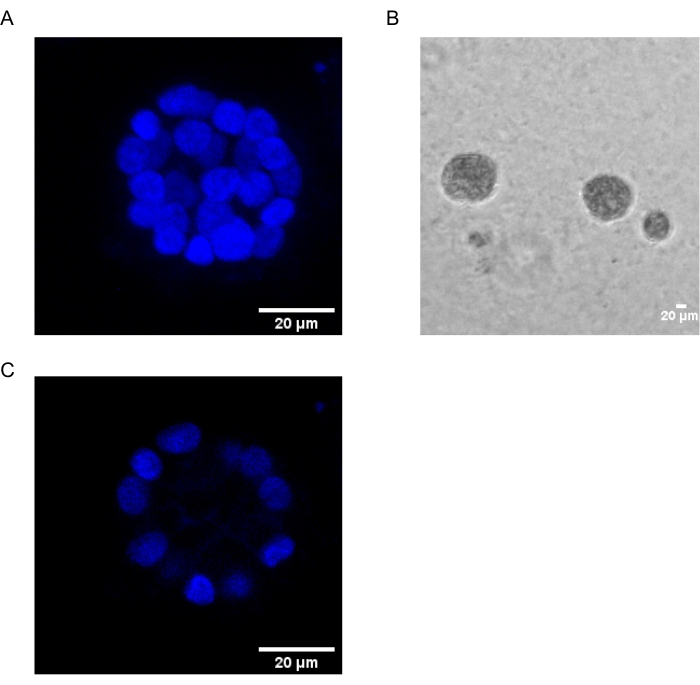
Figura 1: Núcleos de células en acinos teñidos con una tinción nuclear . (A) Construcción 3D de los acinos. (B) Imagen de contraste de fase de acinos MCF10A cultivados en Matrigel durante 20 días. (C) La sección central muestra la presencia de un lumen hueco. Barra de escala = 20 μm. Haga clic aquí para ver una versión más grande de esta figura.
A diferencia de los cultivos 2D, los cultivos acinares ayudan a distinguir las células normales y transformadas a través de cambios morfológicos aparentes. Las células epiteliales mamarias no transformadas forman acinos con una luz hueca, imitando los acinos normales de la mama humana. Estos esferoides, tras la transformación, muestran una morfología alterada caracterizada por una pérdida importante de polaridad (una de las características del cáncer), ausencia de una luz o interrupción de la luz hueca (debido a la evasión de la apoptosis) que puede ser inducida debido a la desregulación de varios genes17,18,19,20 . Estas transformaciones se pueden estudiar utilizando técnicas de uso común como la inmunofluorescencia. Por lo tanto, el modelo de cultivo celular 3D puede funcionar como un método simple para investigar el proceso de morfogénesis acinar mamaria y carcinogénesis mamaria. El establecimiento de un sistema de cultivo 3D para comprender el efecto de un mediador fosfolípido, PAF, ayudará en la detección preclínica de fármacos de alto rendimiento.
Este trabajo ha adaptado el protocolo de cultivo 3D ‘on top’16,21 para estudiar la transformación inducida por PAF 22. Los cambios fenotípicos inducidos por la exposición de los acinos al mediador fosfolípido se estudiaron mediante inmunofluorescencia. En el estudio se utilizaron varios marcadores de polaridad y transición epitelial-mesenquimal (EMT)12,16. La Tabla 1 menciona su localización normal y su fenotipo esperado tras la transformación.
| Anticuerpos | Marcas | Localización normal | Fenotipo transformado |
| α6-Integrina | Basolateral | Basal con tinción lateral débil | Fuerte tinción lateral / apical |
| β-Catenin | Unión célula-célula | Basolateral | Localización anormal / nuclear o citoplasmática |
| Vimentina | Emt | Presencia ausente / débil | Regulación positiva |
Tabla 1: Marcadores utilizados en el estudio. Diferentes marcadores utilizados con su localización en presencia y ausencia de tratamiento con FAP.
Este método se puede utilizar mejor para estudiar / detectar medicamentos plausibles y genes objetivo para varios subtipos de cáncer de mama. Esto puede proporcionar datos de respuesta a medicamentos más cercanos al escenario in vivo , ayudando a un desarrollo de medicamentos más rápido y confiable. Además, este sistema se puede utilizar para estudiar la señalización molecular asociada con la respuesta a los medicamentos y la resistencia a los medicamentos.
