מספר עצום של מודלים זמינים כדי לחקור את התקדמות הסרטן, כל אחד מהם להיות ייחודי ומייצג תת סוג של מחלה מורכבת זו. כל מודל מספק תובנות ייחודיות ובעלות ערך על הביולוגיה של הסרטן ושיפר את האמצעים לחקות את מצב המחלה בפועל. קווי תאים מבוססים שגודלו כחד-שכבתיים סיפקו תובנות חשובות על תהליכים חיוניים במבחנה, כגון התפשטות, פולשות, נדידה ואפופטוזיס1. אף על פי שתרבית תאים דו-ממדית (2D) היא הכלי המסורתי לחקור את התגובה של תאי יונקים למספר הפרעות סביבתיות, אקסטרפולציה של ממצאים אלה כדי לחזות תגובות ברמת הרקמה אינה נראית משכנעת מספיק. המגבלה העיקרית של התרביות הדו-ממדיות היא שהמיקרו-סביבה שנוצרת שונה במידה רבה מזו של רקמת השד עצמה2. תרבית דו-ממדית חסרה את האינטראקציה של התאים עם המטריצה החוץ-תאית, החיונית לצמיחה של כל רקמה. כמו כן, כוחות מתיחה שחווה התא בתרביות חד-שכבתיות מעכבים את הקוטביות של תאים אלה, ובכך משנים את האיתות וההתנהגות של התא 3,4,5. מערכות תרבית תלת-ממדיות (תלת-ממדיות) פתחו אפיק חדש בתחום חקר הסרטן עם יכולתן לחקות את תנאי ה-in vivo במבחנה. רמזים מיקרו-סביבתיים חיוניים רבים שאבדו בתרבית תאים דו-ממדית יכולים להתבסס מחדש באמצעות תרביות תלת-ממדיות של מטריצה חוץ-תאית עשירה בלמינין (lrECM)6.
מחקרים שונים זיהו את החשיבות של מיקרו-סביבה של הגידול בסרטן 7,8. גורמים הקשורים לדלקת הם חלק מרכזי מהמיקרו-סביבה. גורם מפעיל טסיות (PAF) הוא מתווך פוספוליפידים המופרש על ידי תאי חיסון שונים המתווך תגובות חיסוניות מרובות 9,10. רמות גבוהות של PAF מופרשות על ידי קווי תאים שונים של סרטן השד וקשורות לשגשוג מוגבר11. מחקרים מהמעבדה שלנו הראו כי נוכחות ממושכת של PAF בתרביות אצנר מובילה לשינוי של תאי אפיתל השד12. PAF מפעיל את קולטן ה-PAF (PAFR), ומפעיל את ציר האיתות PI3K/Akt13. דווח כי PAFR קשור גם ל-EMT, פלישה וגרורות14.
הפרוטוקול הנוכחי מדגים מערכת מודל לחקר טרנספורמציה המושרה על ידי PAF, תוך שימוש בתרביות תלת-ממדיות של תאי אפיתל השד, כפי שתואר בעבר על ידי Chakravarty et al.12. תאי אפיתל השד הגדלים על המטריצה החוץ-תאית (תרביות תלת-ממדיות) נוטים ליצור ספרואידים מקוטבים שנעצרו על-ידי צמיחה. אלה נקראים acini ודומים מאוד acini של רקמת השד, היחידה הפונקציונלית הקטנה ביותר של בלוטת החלב, in vivo15. הספרואידים האלה (איור 1A,B) מורכבים ממונו-שכבה של תאי אפיתל מקוטבים צפופים שמקיפים לומן חלול ומחוברים לקרום המרתף (איור 1C). תהליך זה של מורפוגנזה תואר היטב בספרות16. כאשר הם נזרעים על lrECM, התאים עוברים חלוקה והתמיינות כדי ליצור אשכול תאים, אשר לאחר מכן מקטבים מיום 4 ואילך. עד יום 8, האצ’יני מורכב מקבוצה של תאים מקוטבים הנמצאים במגע ישיר עם המטריצה החוץ-תאית וצביר של תאים לא מקוטבים הסגורים בתוך התאים המקוטבים החיצוניים, ללא מגע עם המטריצה. ידוע כי תאים לא מקוטבים אלה עוברים אפופטוזיס עד יום 12 של התרבית, ויוצרים לומן חלול. עד יום 16, מבנים שנעצרו על ידי צמיחה נוצרים16.
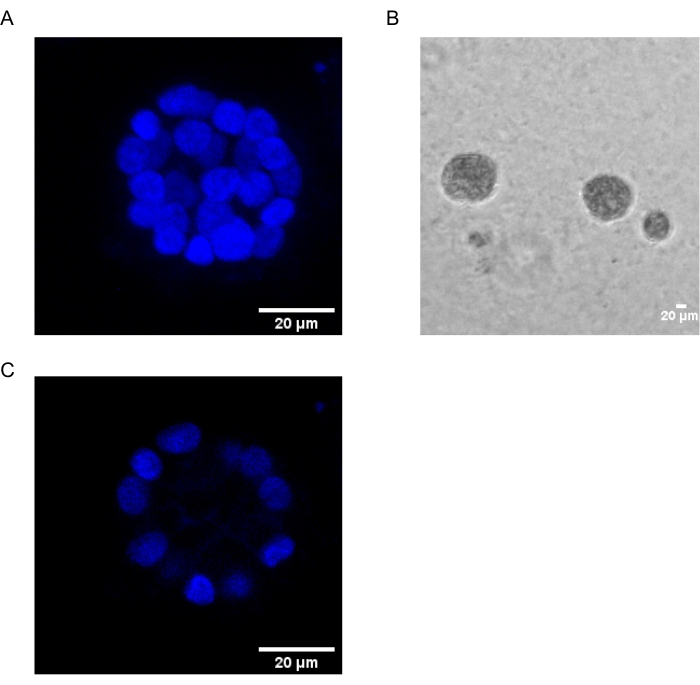
איור 1: גרעינים של תאים באצ’יני המוכתמים בכתם גרעיני . (A) בנייה תלת-ממדית של האצ’יני. (B) תמונת ניגודיות פאזה של MCF10A acini שגדלה על Matrigel במשך 20 יום. (C) החלק המרכזי ביותר מראה נוכחות של לומן חלול. סרגל קנה מידה = 20 מיקרומטר. אנא לחץ כאן כדי להציג גרסה גדולה יותר של נתון זה.
שלא כמו תרביות דו-ממדיות, תרביות אצ’ינאר מסייעות להבחין בין תאים נורמליים לתאים שעברו טרנספורמציה באמצעות שינויים מורפולוגיים לכאורה. תאי אפיתל שד שאינם מותמרים יוצרים acini עם לומן חלול, המחקה את אציני השד האנושי הרגיל. ספרואידים אלה, עם הטרנספורמציה, מראים מורפולוגיה משובשת המאופיינת באובדן גדול של קוטביות (אחד מסימני ההיכר של סרטן), היעדר לומן, או שיבוש של לומן חלול (עקב התחמקות מאפופטוזיס) שעלול להיגרם עקב הסרת רגולציה של גנים שונים17,18,19,20 . ניתן לחקור טרנספורמציות אלה באמצעות טכניקות נפוצות כגון אימונופלואורסצנציה. לפיכך, מודל תרבית התאים התלת-ממדית יכול לתפקד כשיטה פשוטה לחקור את התהליך של מורפוגנזה של אצנר השד וסרטן השד. הקמת מערכת תרבית תלת-ממדית להבנת ההשפעה של מתווך פוספוליפידים, PAF, תסייע בבדיקת תרופות פרה-קלינית בתפוקה גבוהה.
עבודה זו התאימה את פרוטוקול התרבות התלת-ממדית ‘על גבי‘ 16,21 לחקר טרנספורמציה המושרה על ידי PAF 22. השינויים הפנוטיפיים הנגרמים על ידי חשיפה של האציני למתווך הפוספוליפידים נחקרו באמצעות אימונופלואורסצנציה. במחקר נעשה שימוש בסמני קוטביות ואפיתל שונים למעבר מזנכימלי (EMT). טבלה 1 מזכירה את הלוקליזציה הרגילה שלהם ואת הפנוטיפ הצפוי שלהם בעת הטרנספורמציה.
| נוגדנים | סימני | לוקליזציה רגילה | פנוטיפ מותמר |
| α6-אינטגרין | בזולטרלי | בזל עם כתם רוחבי חלש | כתם רוחבי / אפיקלי חזק |
| β-קטנין | צומת תא-תא | בזולטרלי | לוקליזציה חריגה / גרעינית או ציטופלסמית |
| וימנטין | EMT | נוכחות נעדרת / חלשה | אפ-רגולציה |
טבלה 1: סמנים ששימשו במחקר. סמנים שונים המשמשים עם לוקליזציה שלהם בנוכחות והיעדר טיפול PAF.
ניתן להשתמש בשיטה זו בצורה הטובה ביותר כדי לחקור/לסנן תרופות סבירות ולמקד גנים עבור תתי-סוגים שונים של סרטן השד. זה יכול לספק נתוני תגובה לתרופות קרוב יותר לתרחיש in vivo , ולסייע בפיתוח תרופות מהיר ואמין יותר. כמו כן, ניתן להשתמש במערכת זו כדי לחקור את האיתות המולקולרי הקשור לתגובה לתרופות ולעמידות לתרופות.
