Для изучения прогрессирования рака доступно множество моделей, каждая из которых уникальна и представляет собой подтип этого сложного заболевания. Каждая модель предоставляет уникальную и ценную информацию о биологии рака и улучшает средства для имитации фактического состояния заболевания. Установленные клеточные линии, выращенные как монослой, предоставили ценную информацию о жизненно важных процессах in vitro, таких как пролиферация, инвазивность, миграция и апоптоз1. Хотя двумерная (2D) клеточная культура была традиционным инструментом для исследования реакции клеток млекопитающих на несколько возмущений окружающей среды, экстраполяция этих результатов для прогнозирования реакций на тканевом уровне не кажется достаточно убедительной. Основным ограничением 2D-культур является то, что создаваемая микросреда в значительной степени отличается от микросреды самой ткани молочной железы2. 2D культуре не хватает взаимодействия клеток с внеклеточным матриксом, что жизненно важно для роста любой ткани. Кроме того, силы растяжения, испытываемые клеткой в монослойных культурах, препятствуют полярности этих клеток, тем самым изменяя передачу сигналов и поведение клеток 3,4,5. Трехмерные (3D) системы культур открыли новый путь в области исследований рака с их способностью имитировать условия in vivo in vitro. Многие важные микроокружательные сигналы, которые теряются в 2D-культуре клеток, могут быть восстановлены с использованием 3D-культур богатого ламинином внеклеточного матрикса (lrECM)6.
Различные исследования выявили значение микроокружения опухоли в канцерогенезе 7,8. Факторы, связанные с воспалением, являются основной частью микросреды. Активирующий фактор тромбоцитов (PAF) является фосфолипидным медиатором, секретируемым различными иммунными клетками, который опосредует множественные иммунные реакции 9,10. Высокие уровни PAF секретируются различными клеточными линиями рака молочной железы и связаны с усиленной пролиферацией11. Исследования из нашей лаборатории показали, что длительное присутствие PAF в ацинарных культурах приводит к трансформации эпителиальных клеток молочной железы12. PAF активирует PAF-рецептор (PAFR), активируя сигнальную ось13 PI3K/Akt. Также сообщается, что PAFR связан с EMT, инвазией и метастазированием14.
Настоящий протокол демонстрирует модельную систему для изучения PAF-индуцированной трансформации с использованием 3D-культур эпителиальных клеток молочной железы, как было ранее описано Chakravarty et al.12. Эпителиальные клетки молочной железы, выращенные на внеклеточном матриксе (3D-культуры), имеют тенденцию образовывать поляризованные сфероиды, остановленные ростом. Они называются ацинами и очень напоминают ацины ткани молочной железы, наименьшую функциональную единицу молочной железы, in vivo15. Эти сфероиды (фиг.1А,В) состоят из монослоя плотно упакованных поляризованных эпителиальных клеток, окружающих полый просвет и прикрепленных к базальной мембране (фиг.1С). Этот процесс морфогенеза хорошо описан в литературе16. При посеве на lrECM клетки подвергаются делению и дифференцировке, образуя кластер клеток, которые затем поляризуются с 4-го дня. К 8-му дню ацины состоят из группы поляризованных клеток, которые находятся в прямом контакте с внеклеточным матриксом и скопления неполяризованных клеток, заключенных во внешние поляризованные клетки, без контакта с матрицей. Известно, что эти неполяризованные клетки подвергаются апоптозу на 12-й день культуры, образуя полый просвет. К 16-му дню формируется16 структур, остановленных ростом.
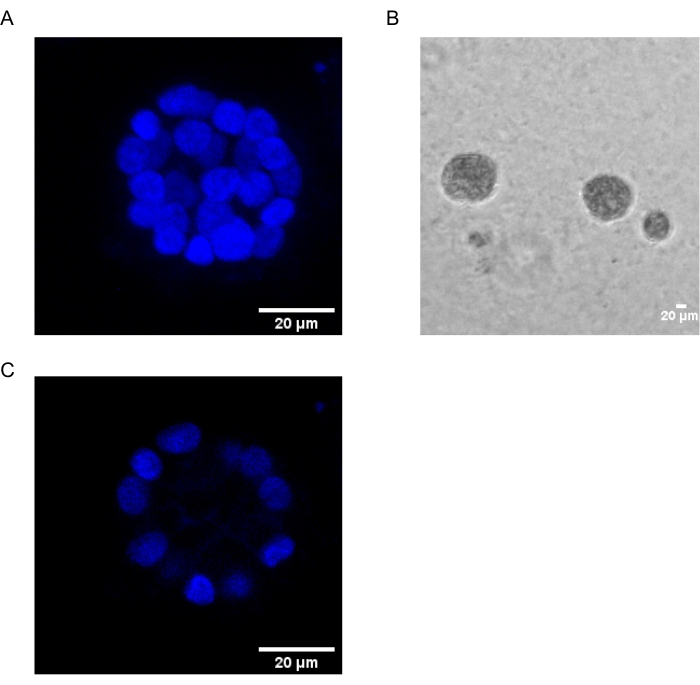
Рисунок 1: Ядра клеток в ацини, окрашенные ядерным пятном. (А) 3D конструкция ацини. (B) Фазоконтрастное изображение ацини MCF10A, выращенное на Matrigel в течение 20 дней. (C) Самая центральная секция показывает наличие полого просвета. Шкала = 20 мкм. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
В отличие от 2D-культур, ацинарные культуры помогают различать нормальные и трансформированные клетки через очевидные изменения морфологии. Нетрансформированные эпителиальные клетки молочной железы образуют ацинус с полым просветом, имитируя нормальные ацины молочной железы человека. Эти сфероиды при трансформации показывают нарушенную морфологию, характеризующуюся значительной потерей полярности (одним из признаков рака), отсутствием просвета или нарушением полого просвета (из-за уклонения от апоптоза), что может быть вызвано дерегуляцией различных генов 17,18,19,20 . Эти превращения могут быть изучены с использованием широко используемых методов, таких как иммунофлуоресценция. Таким образом, модель 3D-культуры клеток может функционировать как простой метод исследования процесса ацинарного морфогенеза молочной железы и канцерогенеза молочной железы. Создание системы 3D-культур для понимания эффекта медиатора фосфолипидов, PAF, поможет в высокопроизводительном доклиническом скрининге лекарств.
Эта работа адаптировала протокол 3D «на вершине» культуры 16,21 для изучения трансформации, вызванной PAF22. Фенотипические изменения, вызванные воздействием ацинумов на медиатор фосфолипидов, изучали с использованием иммунофлуоресценции. В исследовании использовались различные маркеры полярности и эпителиального перехода в мезенхимальный переход (ЭМТ) 12,16. В таблице 1 упоминается их нормальная локализация и ожидаемый фенотип при трансформации.
| Антитела | Знаки | Нормальная локализация | Трансформированный фенотип |
| α6-Интегрин | Базолатераль | Базальный со слабым боковым пятном | Сильное боковое / апикальное пятно |
| β-Катенин | Клеточно-клеточный переход | Базолатераль | Аномальная / ядерная или цитоплазматическая локализация |
| Виментин | ЭМТ | Отсутствующее/слабое присутствие | Ап-регулирование |
Таблица 1: Маркеры, использованные в исследовании. Используются различные маркеры с их локализацией при наличии и отсутствии лечения ПАФ.
Этот метод может быть наилучшим образом использован для изучения / скрининга правдоподобных лекарств и генов-мишеней для различных подтипов рака молочной железы. Это может обеспечить данные о реакции на лекарства ближе к сценарию in vivo , помогая в более быстрой и надежной разработке лекарств. Также эта система может быть использована для изучения молекулярной сигнализации, связанной с лекарственной реакцией и лекарственной устойчивостью.
