En myriad av modeller finns tillgängliga för att studera utvecklingen av cancer, var och en av dem är unik och representerar en subtyp av denna komplexa sjukdom. Varje modell ger unika och värdefulla insikter i cancerbiologi och har förbättrat medlen för att efterlikna det faktiska sjukdomstillståndet. Etablerade cellinjer som odlas som ett monolager har gett värdefulla insikter i viktiga processer in vitro, såsom spridning, invasivitet, migration och apoptos1. Även om tvådimensionell (2D) cellodling har varit det traditionella verktyget för att undersöka däggdjurscellernas svar på flera miljöstörningar, verkar extrapolering av dessa fynd för att förutsäga vävnadsnivåsvar inte tillräckligt övertygande. Den största begränsningen av 2D-kulturerna är att den skapade mikromiljön skiljer sig till stor del från bröstvävnaden i sig2. 2D-kulturen saknar interaktionen mellan cellerna och den extracellulära matrisen, vilket är avgörande för tillväxten av vilken vävnad som helst. Dragkrafter som upplevs av cellen i monolagerkulturer hindrar också polariteten hos dessa celler, vilket förändrar cellsignalering och beteende 3,4,5. Tredimensionella (3D) odlingssystem har öppnat en ny väg inom cancerforskningen med sin förmåga att efterlikna in vivo-förhållandena in vitro. Många viktiga mikromiljösignaler som går förlorade i 2D-cellodling kan återupprättas med hjälp av 3D-kulturer av lamininrik extracellulär matris (lrECM)6.
Olika studier har identifierat betydelsen av tumörmikromiljön vid cancerframkallande 7,8. Inflammationsassocierade faktorer är en stor del av mikromiljön. Trombocytaktiveringsfaktor (PAF) är en fosfolipidmediator som utsöndras av olika immunceller som förmedlar flera immunsvar 9,10. Höga nivåer av PAF utsöndras av olika bröstcancercellinjer och är förknippade med förbättrad proliferation11. Studier från vårt laboratorium har visat att den långvariga närvaron av PAF i acinarkulturer leder till omvandling av bröstepitelceller12. PAF aktiverar PAF-receptorn (PAFR) och aktiverar PI3K/Akt-signalaxeln13. PAFR rapporteras också vara associerad med EMT, invasion och metastasering14.
Detta protokoll visar ett modellsystem för att studera PAF-inducerad transformation, med hjälp av 3D-kulturer av bröstepitelceller, som tidigare har beskrivits av Chakravarty et al.12. Bröstepitelcellerna som odlas på den extracellulära matrisen (3D-kulturer) tenderar att bilda polariserade tillväxtarresterade sfäroider. Dessa kallas acini och liknar acini i bröstvävnad, den minsta funktionella enheten i bröstkörteln, in vivo15. Dessa sfäroider (figur 1A,B) består av ett monolager av tätt packade polariserade epitelceller som omger en ihålig lumen och är fästa vid källarmembranet (figur 1C). Denna process av morfogenes har beskrivits väl i litteratur16. När de seedas på lrECM genomgår cellerna delning och differentiering för att bilda ett kluster av celler, som sedan polariseras från dag 4 och framåt. Vid dag 8 består acini av en grupp polariserade celler som är i direkt kontakt med den extracellulära matrisen och ett kluster av opolariserade celler inneslutna i de yttre polariserade cellerna, utan kontakt med matrisen. Dessa opolariserade celler är kända för att genomgå apoptos vid dag 12 av kulturen och bildar en ihålig lumen. Vid dag 16 bildas tillväxtarresterade strukturer16.
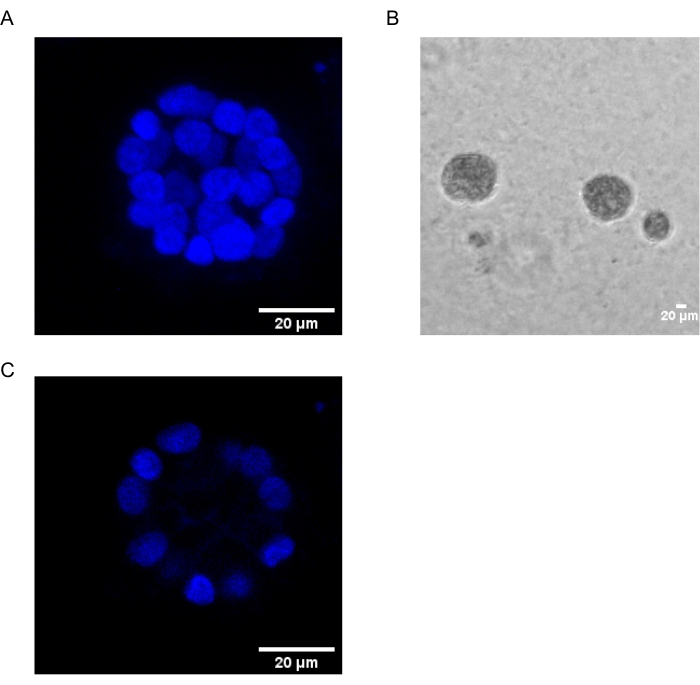
Figur 1: Kärnor av celler i acini färgade med en kärnfläck . (A) 3D-konstruktion av acini. (B) Faskontrastbild av MCF10A acini odlad på Matrigel i 20 dagar. (C) Den mittersta sektionen visar närvaron av en ihålig lumen. Skalstreck = 20 μm. Klicka här för att se en större version av denna figur.
Till skillnad från 2D-kulturer hjälper acinarkulturer att skilja normala och transformerade celler genom uppenbara morfologiska förändringar. Icke-transformerade bröstepitelceller bildar acini med en ihålig lumen som efterliknar den normala mänskliga bröstacini. Dessa sfäroider, vid transformation, visar en störd morfologi som kännetecknas av en stor förlust av polaritet (ett av kännetecknen för cancer), frånvaro av lumen eller störning av den ihåliga lumen (på grund av undvikande av apoptos) som kan induceras på grund av avreglering av olika gener17,18,19,20 . Dessa transformationer kan studeras med hjälp av vanliga tekniker såsom immunofluorescens. Således kan 3D-cellodlingsmodellen fungera som en enkel metod för att undersöka processen för bröstacinarmorfogenes och bröstcancer. Att etablera ett 3D-odlingssystem för att förstå effekten av en fosfolipidmediator, PAF, kommer att hjälpa till med preklinisk läkemedelsscreening med hög genomströmning.
Detta arbete har anpassat 3D ‘on top’ kulturprotokoll16,21 för att studera transformation inducerad av PAF 22. De fenotypiska förändringarna som inducerades av exponering av acini för fosfolipidmediatorn studerades med användning av immunofluorescens. Olika markörer för polaritet och epitelial till mesenkymal övergång (EMT)12,16 användes i studien. Tabell 1 nämner deras normala lokalisering och deras förväntade fenotyp vid transformation.
| Antikroppar | Markerar | Normal lokalisering | Transformerad fenotyp |
| α6-Integrin | Basolateral | Basal med svag lateral fläck | Stark lateral / apikal fläck |
| β-Catenin | Korsning mellan cell och cell | Basolateral | Onormal / nukleär eller cytoplasmatisk lokalisering |
| Vimentin | Emt | Frånvarande / svag närvaro | Uppreglering |
Tabell 1: Markörer som använts i studien. Olika markörer som används med deras lokalisering i närvaro och frånvaro av PAF-behandling.
Denna metod kan bäst användas för att studera / screena troliga läkemedel och målgener för olika bröstcancerundertyper. Detta kan ge en läkemedelsresponsdata närmare in vivo-scenariot , vilket hjälper till med snabbare och mer tillförlitlig läkemedelsutveckling. Detta system kan också användas för att studera molekylär signalering associerad med läkemedelssvar och läkemedelsresistens.
