In 2011, researchers proposed a computational method and developed a corresponding piece of software for the automatic design of digital synthetic gene circuits1. A user had to specify the number of inputs (three or four) and fill in the circuit truth table; this provided all the necessary information to derive the circuit structure using techniques from electronics. The truth table was translated into two Boolean formulae via the Karnaugh map method2. Each Boolean formula is made of clauses that describe logic operations (sum or multiplication) among (part of) the circuit inputs and their negations (the literals). Clauses, in their turn, are either summed up (OR) or multiplied (AND) to compute the circuit output. Every circuit can be realized according to any of its two corresponding formulae: one written in POS (product of sums) form and the other in SOP (sum of products) representation. The former consists of a multiplication of clauses (i.e., Boolean gates) that contain a logic sum of the literals. The latter, in contrast, is a sum of clauses where the literals are multiplied.
Electric circuits can be realized, on a breadboard, by physically wiring different gates together. The electric current permits the exchange of signals among gates, which leads to the computation of the output.
In biology, the situation is more complex. A Boolean gate can be realized as a transcription unit (TU; i.e., the sequence "promoter-coding region-terminator" inside eukaryotic cells), where transcription or translation (or both) are regulated. Thus, at least two kinds of molecules establish a biological wiring: the transcription factor proteins and the non-coding, antisense RNAs1.
A gene digital circuit is organized into two or three layers of gates, namely: 1) the input layer, which is made of YES (buffer) and NOT gates and converts the input chemicals into wiring molecules; 2) the internal layer, which consists of as many TUs as there are clauses in the corresponding Boolean formula. If the circuit is designed according to the SOP formula, every clause in the internal layer will produce the circuit output (e.g., fluorescence) in a so-called distributed output architecture. If the product of sum (POS) formula is used, then a 3) final layer is required, which will contain a single multiplicative gate collecting the wiring molecules from the internal layer.
Overall, in synthetic biology, many different schemes can be designed for the same circuit. They differ in the number and the kind of both TUs and wiring molecules. In order to choose the easiest solution to be implemented in yeast cells, each circuit design is associated with a complexity score S, defined as
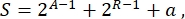
where A represents the number of activators, R represents the number of repressors, and a is the amount of antisense RNA molecules. If either activators or repressors are absent from the circuit, their contribution to S is zero. Therefore, it is more difficult to realize a circuit scheme in the lab (high S) when it requires a high number of orthogonal transcription factors. This means that new activators and repressors shall be engineered de novo in order to realize the complete wiring inside the digital circuits. In principle, novel DNA-binding proteins can be assembled by using Zinc Finger proteins3 and TAL effectors4 as templates. However, this option appears too arduous and time-consuming; therefore, one should rely mostly on small RNAs and translation regulation to finalize complex gene circuits.
Originally, this method was developed to fabricate digital circuits in bacteria. Indeed, in eukaryotic cells, instead of antisense RNAs, it is more suitable to talk of microRNAs (miRNAs) or small interfering RNAs (siRNAs)5. However, the RNAi pathway is not present in the yeast S. cerevisiae. Hence, one should opt for fully transcriptional networks. Suppose that a circuit needs five activators and five repressors; its complexity score would be S = 32. Circuit complexity can be reduced by replacing the 10 transcription factors with a single dCas96 (nuclease deficient Cas9) fused to an activation domain (AD). As shown in7, dCas9-AD works as a repressor in yeast when binding a promoter between the TATA box and the TSS (transcription start site) and as an activator when binding well upstream of the TATA box. Thus, one can replace 10 transcription factors with a single dCas9-AD fusion protein and 10 sgRNAs (single guide RNAs) for a total complexity score of S = 11. It is quick and easy to synthesize ten sgRNAs, whereas, as previously commented, the assembly of 10 proteins would demand much longer and more complicated work.
Alternatively, one might use two orthogonal dCas proteins (e.g., dCas9 and dCas12a): one to fuse to an AD, and the other bare or in combination with a repression domain. The complexity score would increase by only one unit (S = 12). Hence, CRISPR-dCas systems are the key to the construction of very intricate gene digital circuits in S. cerevisiae.
This paper deeply characterizes the efficiency of both dCas9- and dCas12a-based repressors and activators in yeast. Results show that they do not demand a high amount of sgRNA to optimize their activity, so episomal plasmids are preferentially avoided. Moreover, dCas9-based activators are far more effective when using a scaffold RNA (scRNA) that recruits copies of the VP64 AD. In contrast, dCas12a works well when fused to the strong VPR AD directly. Furthermore, a synthetic activated promoter demands a variable number of target sites, depending on the configuration of the activator (e.g., three when using dCas12a-VPR, six for dCas9-VP64, and only one with dCas9 and a scRNA). As a repressor, dCas12a appears more incisive when binding the coding region rather than the promoter.
As a drawback, however, CRISPR-dCas9/dCas12a do not interact with chemicals directly. Therefore, they might be of no use in the input layer. For this reason, alternative Boolean gate designs containing anti-CRISPR proteins (Acrs) have been investigated. Acrs act on (d)Cas proteins and inhibit their working8. Hence, they are a means to modulate the activity of CRISPR-(d)Cas systems. This paper thoroughly analyzes the interactions between type II Acrs and (d)Cas9, as well as type V Acrs and (d)Cas12a in S. cerevisiae. Since Acrs are much smaller than Cas proteins, a NOT gate responsive to the estrogen β-estradiol was built by fusing the hormone-binding domain of the human estrogen receptor9-HBD(hER)-to AcrIIA4. Besides, a handful of YES and NOT gates that expressed dCas12a(-AD) constitutively and AcrVAs upon induction with galactose were realized. At present, these gates serve only as a proof of concept. However, they also represent the first step toward a deep rethinking of the algorithm to carry out the computational automatic design of synthetic gene digital circuits in yeast cells.
