2011年、研究者は計算方法を提案し、デジタル合成遺伝子回路の自動設計に対応するソフトウェアを開発しました1。ユーザーは入力数(3つまたは4つ)を指定し、回路真理値表に記入する必要がありました。これにより、電子機器の技術を使用して回路構造を導き出すために必要なすべての情報が提供されました。真理値表は、Karnaughマップ法2を介して2つのブール式に変換されました。各ブール式は、回路入力(の一部)とその否定(リテラル)間の論理演算(合計または乗算)を記述する句で構成されています。句は、順番に合計(OR)または乗算(AND)されて回路出力を計算します。すべての回路は、POS(合計の積)形式で記述された式とSOP(製品の合計)表現で記述された2つの対応する式のいずれかに従って実現できます。前者は、リテラルの論理和を含む句(つまり、ブールゲート)の乗算で構成されます。対照的に、後者は、リテラルが乗算される句の合計です。
電気回路は、ブレッドボード上で、異なるゲートを物理的に配線することによって実現できます。電流はゲート間の信号交換を可能にし、出力の計算につながります。
生物学では、状況はより複雑です。ブールゲートは、転写または翻訳(またはその両方)が調節される転写単位(TU、すなわち、真核細胞内の配列「プロモーター-コード領域-ターミネーター」)として実現することができる。したがって、少なくとも2種類の分子、すなわち転写因子タンパク質および非コードアンチセンスRNA1の生物学的配線を確立する。
遺伝子デジタル回路は、2層または3層のゲート、すなわち1)YES(バッファー)およびNOTゲートで構成された入力層で構成され、入力化学物質を配線分子に変換する。2)対応するブール式に句があるのと同じ数のTUで構成される内部層。回路がSOP式に従って設計されている場合、内部層のすべての句は、いわゆる分散出力アーキテクチャで回路出力(蛍光など)を生成します。和の積(POS)式が使用される場合、3)最終層が必要であり、これには内部層から配線分子を収集する単一の乗法ゲートが含まれます。
全体として、合成生物学では、同じ回路に対して多くの異なるスキームを設計することができます。それらはTUと配線分子の両方の数と種類が異なります。酵母細胞に実装する最も簡単なソリューションを選択するために、各回路設計は、次のように定義される複雑さスコアSに関連付けられています。
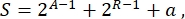
ここで、 A は活性化剤の数を表し、 R はリプレッサーの数を表し、 a はアンチセンスRNA分子の量です。アクティベータまたはリプレッサーのいずれかが回路にない場合、Sへの寄与はゼロです。そのため、ラボでの回路スキーム(高S)の実現は、多数の直交転写因子を必要とする場合により困難である。これは、デジタル回路内の完全な配線を実現するために、新しいアクティベータとリプレッサーを de novo で設計する必要があることを意味します。原理的には、ジンクフィンガータンパク質3 およびTALエフェクター4 を鋳型として使用することにより、新規DNA結合タンパク質を組み立てることができる。ただし、このオプションは非常に困難で時間がかかるように見えます。したがって、複雑な遺伝子回路を完成させるために、主に小さなRNAと翻訳調節に頼る必要があります。
もともと、この方法は細菌のデジタル回路を作製するために開発されました。実際、真核細胞では、アンチセンスRNAの代わりに、マイクロRNA(miRNA)または低分子干渉RNA(siRNA)について話す方が適しています5。しかしながら、RNAi経路は酵母 S.セレビシエには存在しない。したがって、完全に転写ネットワークを選択する必要があります。回路に5つのアクティベータと5つのリプレッサーが必要であるとします。その複雑さのスコアはS = 32になります。10個の転写因子を活性化ドメイン(AD)に融合した単一のdCas96 (ヌクレアーゼ欠損Cas9)に置き換えることで、回路の複雑さを軽減することができます。7に示すように、dCas9-ADは、TATAボックスとTSS(転写開始部位)の間でプロモーターを結合させる際の酵母のリプレッサーとして、またTATAボックスのかなり上流に結合するときの活性化剤として働きます。したがって、10個の転写因子を単一のdCas9-AD融合タンパク質と10個のsgRNA(シングルガイドRNA)に置き換えることができ、合計複雑さスコアはS = 11になります。10個のsgRNAを合成するのは迅速かつ簡単ですが、前述のように、10個のタンパク質の組み立てにははるかに長く複雑な作業が必要になります。
あるいは、2つの直交するdCasタンパク質(例えば、dCas9およびdCas12a)を使用することができ、1つはADに融合し、もう1つは裸または抑制ドメインと組み合わせて使用する。複雑さのスコアは 1 単位だけ増加します (S = 12)。したがって、CRISPR-dCasシステムは、 出芽酵母における非常に複雑な遺伝子デジタル回路の構築の鍵となります。
この論文は、酵母におけるdCas9およびdCas12aベースのリプレッサーおよびアクチベーターの両方の効率を深く特徴付けています。結果は、それらがそれらの活性を最適化するために大量のsgRNAを必要としないことを示したので、エピソームプラスミドは優先的に避けられる。さらに、dCas9ベースのアクチベーターは、VP64 ADのコピーをリクルートするスキャフォールドRNA(scRNA)を使用する場合にはるかに効果的です。対照的に、dCas12aは、強力なVPR ADに直接融合するとうまく機能します。さらに、合成活性化プロモーターは、活性化因子の構成に応じて、可変数の標的部位を必要とします(例えば、dCas12a-VPRを使用する場合は3つ、dCas9-VP64を使用する場合は6つ、dCas9とscRNAを使用する場合は1つだけ)。リプレッサーとして、dCas12aはプロモーターよりもコード領域に結合するとより鋭敏に見えます。
ただし、欠点として、CRISPR-dCas9 / dCas12aは化学物質と直接相互作用しません。したがって、入力層では役に立たない可能性があります。このため、抗CRISPRタンパク質(ACRS)を含む代替ブールゲート設計が検討されています。ACRは(d)Casタンパク質に作用し、その働きを阻害します8.したがって、それらはCRISPR-(d)Casシステムの活性を調節する手段です。この論文では、出芽酵母のII型ACRSと(d)Cas9、およびV型Acrと(d)Cas12aの相互作用を徹底的に分析しています。AcrsはCasタンパク質よりもはるかに小さいため、ヒトエストロゲン受容体9-HBD(hER)-のホルモン結合ドメインをAcrIIA4に融合させることによって、エストロゲンβ-エストラジオールに応答するNOTゲートを構築しました。さらに、dCas12a(-AD)を構成的に発現する少数のYESゲートとNOTゲート、およびガラクトースによる誘導時にAcrVAが実現されました。現在、これらのゲートは概念実証としてのみ機能します。しかし、それらはまた、酵母細胞における合成遺伝子デジタル回路の計算自動設計を実行するためのアルゴリズムの深い再考に向けた第一歩を表しています。
