En 2011, des chercheurs ont proposé une méthode de calcul et développé un logiciel correspondant pour la conception automatique de circuits génétiques synthétiques numériques1. Un utilisateur devait spécifier le nombre d’entrées (trois ou quatre) et remplir la table de vérité du circuit; Cela a fourni toutes les informations nécessaires pour dériver la structure du circuit en utilisant des techniques de l’électronique. La table de vérité a été traduite en deux formules booléennes via la méthode2 de la carte de Karnaugh. Chaque formule booléenne est constituée de clauses qui décrivent des opérations logiques (somme ou multiplication) entre (partie de) les entrées du circuit et leurs négations (les littéraux). Les clauses, à leur tour, sont soit additionnées (OR) ou multipliées (ET) pour calculer la sortie du circuit. Chaque circuit peut être réalisé selon l’une de ses deux formules correspondantes : l’une écrite sous forme POS (produit des sommes) et l’autre en SOP (somme des produits). Le premier consiste en une multiplication de clauses (c’est-à-dire des portes booléennes) qui contiennent une somme logique des littéraux. Ce dernier, en revanche, est une somme de clauses où les littéraux sont multipliés.
Les circuits électriques peuvent être réalisés, sur une planche à pain, en câblant physiquement différentes portes ensemble. Le courant électrique permet l’échange de signaux entre les portes, ce qui conduit au calcul de la sortie.
En biologie, la situation est plus complexe. Une porte booléenne peut être réalisée comme une unité de transcription (TU; c’est-à-dire la séquence « promoteur-codant région-terminateur » à l’intérieur des cellules eucaryotes), où la transcription ou la traduction (ou les deux) sont régulées. Ainsi, au moins deux sortes de molécules établissent un câblage biologique : les protéines du facteur de transcription et les ARN antisens non codants1.
Un circuit numérique génétique est organisé en deux ou trois couches de portes, à savoir: 1) la couche d’entrée, qui est faite de portes YES (tampon) et NOT et convertit les produits chimiques d’entrée en molécules de câblage; 2) la couche interne, qui se compose d’autant de TU qu’il y a de clauses dans la formule booléenne correspondante. Si le circuit est conçu selon la formule SOP, chaque clause de la couche interne produira la sortie du circuit (par exemple, la fluorescence) dans une architecture dite de sortie distribuée. Si la formule du produit de somme (POS) est utilisée, alors une couche finale 3) est nécessaire, qui contiendra une seule porte multiplicative collectant les molécules de câblage de la couche interne.
Dans l’ensemble, en biologie synthétique, de nombreux schémas différents peuvent être conçus pour le même circuit. Ils diffèrent par le nombre et le type de TU et de molécules de câblage. Afin de choisir la solution la plus simple à mettre en œuvre dans les cellules de levure, chaque conception de circuit est associée à un score de complexité S, défini par
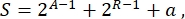
où A représente le nombre d’activateurs, R représente le nombre de répresseurs et a est la quantité de molécules d’ARN antisens. Si les activateurs ou les répresseurs sont absents du circuit, leur contribution à S est nulle. Par conséquent, il est plus difficile de réaliser un schéma de circuit en laboratoire (S élevé) lorsqu’il nécessite un nombre élevé de facteurs de transcription orthogonaux. Cela signifie que de nouveaux activateurs et répresseurs doivent être conçus de novo afin de réaliser le câblage complet à l’intérieur des circuits numériques. En principe, de nouvelles protéines de liaison à l’ADN peuvent être assemblées en utilisant les protéines Zinc Finger3 et les effecteurs TAL4 comme modèles. Cependant, cette option semble trop ardue et prend trop de temps; par conséquent, il faut s’appuyer principalement sur les petits ARN et la régulation de la traduction pour finaliser des circuits génétiques complexes.
À l’origine, cette méthode a été développée pour fabriquer des circuits numériques dans des bactéries. En effet, dans les cellules eucaryotes, au lieu d’ARN antisens, il est plus approprié de parler de microARN (miARN) ou de petits ARN interférents (siRNAs)5. Cependant, la voie ARNi n’est pas présente dans la levure S. cerevisiae. Par conséquent, il faut opter pour des réseaux entièrement transcriptionnels. Supposons qu’un circuit ait besoin de cinq activateurs et de cinq répresseurs; son score de complexité serait S = 32. La complexité du circuit peut être réduite en remplaçant les 10 facteurs de transcription par un seul dCas96 (Cas9 déficient en nucléase) fusionné à un domaine d’activation (AD). Comme le montrele point 7, dCas9-AD fonctionne comme un répresseur dans la levure lors de la liaison d’un promoteur entre la boîte TATA et le TSS (site de début de transcription) et comme activateur lors de la liaison bien en amont de la boîte TATA. Ainsi, on peut remplacer 10 facteurs de transcription par une seule protéine de fusion dCas9-AD et 10 sgRNA (ARN guides uniques) pour un score de complexité totale de S = 11. Il est rapide et facile de synthétiser dix ARNg, alors que, comme indiqué précédemment, l’assemblage de 10 protéines nécessiterait un travail beaucoup plus long et plus compliqué.
Alternativement, on peut utiliser deux protéines orthogonales dCas (par exemple, dCas9 et dCas12a): l’une pour fusionner avec une AD, et l’autre nue ou en combinaison avec un domaine de répression. Le score de complexité n’augmenterait que d’une unité (S = 12). Par conséquent, les systèmes CRISPR-dCas sont la clé de la construction de circuits numériques de gènes très complexes chez S. cerevisiae.
Cet article caractérise en profondeur l’efficacité des répresseurs et des activateurs à base de dCas9 et dCas12a chez la levure. Les résultats montrent qu’ils ne nécessitent pas une grande quantité d’ARNg pour optimiser leur activité, de sorte que les plasmides épisomiques sont préférentiellement évités. De plus, les activateurs basés sur dCas9 sont beaucoup plus efficaces lorsqu’ils utilisent un ARN d’échafaudage (scRNA) qui recrute des copies du VP64 AD. En revanche, dCas12a fonctionne bien lorsqu’il est fusionné directement avec le puissant VPR AD. De plus, un promoteur activé synthétique exige un nombre variable de sites cibles, en fonction de la configuration de l’activateur (par exemple, trois lors de l’utilisation de dCas12a-VPR, six pour dCas9-VP64 et un seul avec dCas9 et un scRNA). En tant que répresseur, dCas12a apparaît plus incisif lors de la liaison de la région de codage plutôt que du promoteur.
Cependant, CRISPR-dCas9 / dCas12a n’interagissent pas directement avec les produits chimiques. Par conséquent, ils peuvent ne pas être utiles dans la couche d’entrée. Pour cette raison, d’autres conceptions de portes booléennes contenant des protéines anti-CRISPR (Acrs) ont été étudiées. Les ACRS agissent sur les protéines (d)Cas et inhibent leur fonctionnement8. Par conséquent, ils sont un moyen de moduler l’activité des systèmes CRISPR-(d)Cas. Cet article analyse en profondeur les interactions entre les Acrs de type II et (d)Cas9, ainsi que les Acrs de type V et (d)Cas12a chez S. cerevisiae. Étant donné que les Acr sont beaucoup plus petits que les protéines Cas, une porte NOT sensible à l’œstrogène β-estradiol a été construite en fusionnant le domaine de liaison hormonale du récepteur humaindes œstrogènes 9-HBD(hER)-à AcrIIA4. En outre, une poignée de portes YES et NOT qui exprimaient dCas12a (-AD) de manière constitutive et AcrVAs lors de l’induction avec du galactose ont été réalisées. À l’heure actuelle, ces portes ne servent que de preuve de concept. Cependant, ils représentent également la première étape vers une refonte profonde de l’algorithme pour réaliser la conception automatique computationnelle de circuits numériques de gènes synthétiques dans les cellules de levure.
