Nel 2011, i ricercatori hanno proposto un metodo computazionale e sviluppato un software corrispondente per la progettazione automatica di circuiti genetici sintetici digitali1. Un utente doveva specificare il numero di ingressi (tre o quattro) e compilare la tabella di verità del circuito; Ciò ha fornito tutte le informazioni necessarie per derivare la struttura del circuito utilizzando tecniche dall’elettronica. La tavola di verità è stata tradotta in due formule booleane tramite il metodo 2 della mappa di Karnaugh. Ogni formula booleana è composta da clausole che descrivono le operazioni logiche (somma o moltiplicazione) tra (parte di) gli ingressi del circuito e le loro negazioni (i letterali). Le clausole, a loro volta, vengono sommate (OR) o moltiplicate (AND) per calcolare l’uscita del circuito. Ogni circuito può essere realizzato secondo una qualsiasi delle sue due formule corrispondenti: una scritta in forma POS (prodotto di somme) e l’altra in rappresentazione SOP (somma di prodotti). Il primo consiste in una moltiplicazione di clausole (cioè porte booleane) che contengono una somma logica dei letterali. Quest’ultimo, al contrario, è una somma di clausole in cui i valori letterali sono moltiplicati.
I circuiti elettrici possono essere realizzati, su una breadboard, collegando fisicamente diversi cancelli insieme. La corrente elettrica consente lo scambio di segnali tra i gate, che porta al calcolo dell’uscita.
In biologia, la situazione è più complessa. Un gate booleano può essere realizzato come unità di trascrizione (TU; cioè la sequenza “promotore-codificante regione-terminatore” all’interno di cellule eucariotiche), dove la trascrizione o la traduzione (o entrambe) sono regolate. Quindi, almeno due tipi di molecole stabiliscono un cablaggio biologico: le proteine del fattore di trascrizione e gli RNA antisenso non codificanti1.
Un circuito digitale genico è organizzato in due o tre strati di porte, vale a dire: 1) lo strato di ingresso, che è costituito da SÌ (buffer) e NON porte e converte le sostanze chimiche in ingresso in molecole di cablaggio; 2) il livello interno, che consiste di tanti TU quante sono le clausole nella corrispondente formula booleana. Se il circuito è progettato secondo la formula SOP, ogni clausola nello strato interno produrrà l’uscita del circuito (ad esempio, fluorescenza) in una cosiddetta architettura di uscita distribuita. Se viene utilizzata la formula del prodotto della somma (POS), è necessario uno strato finale 3), che conterrà un singolo gate moltiplicativo che raccoglie le molecole di cablaggio dallo strato interno.
Nel complesso, in biologia sintetica, molti schemi diversi possono essere progettati per lo stesso circuito. Differiscono nel numero e nel tipo di TU e molecole di cablaggio. Al fine di scegliere la soluzione più semplice da implementare nelle cellule di lievito, ogni progetto di circuito è associato a un punteggio di complessità S, definito come
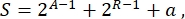
dove A rappresenta il numero di attivatori, R rappresenta il numero di repressori e A è la quantità di molecole di RNA antisenso. Se gli attivatori o i repressori sono assenti dal circuito, il loro contributo a S è zero. Pertanto, è più difficile realizzare uno schema circuitale in laboratorio (alto S) quando richiede un numero elevato di fattori di trascrizione ortogonali. Ciò significa che nuovi attivatori e repressori devono essere progettati de novo per realizzare il cablaggio completo all’interno dei circuiti digitali. In linea di principio, nuove proteine leganti il DNA possono essere assemblate utilizzando le proteine Zinc Finger3 e gli effettori TAL4 come modelli. Tuttavia, questa opzione appare troppo ardua e richiede tempo; pertanto, si dovrebbe fare affidamento principalmente su piccoli RNA e regolazione della traduzione per finalizzare circuiti genici complessi.
Originariamente, questo metodo è stato sviluppato per fabbricare circuiti digitali nei batteri. Infatti, nelle cellule eucariotiche, invece che di RNA antisenso, è più adatto parlare di microRNA (miRNA) o piccoli RNA interferenti (siRNA)5. Tuttavia, la via dell’RNAi non è presente nel lievito S. cerevisiae. Quindi, si dovrebbe optare per reti completamente trascrizionali. Supponiamo che un circuito abbia bisogno di cinque attivatori e cinque repressori; il suo punteggio di complessità sarebbe S = 32. La complessità del circuito può essere ridotta sostituendo i 10 fattori di trascrizione con un singolo dCas96 (Cas9 carente di nucleasi) fuso in un dominio di attivazione (AD). Come mostrato in7, dCas9-AD funziona come repressore nel lievito quando lega un promotore tra la scatola TATA e il TSS (sito di inizio della trascrizione) e come attivatore quando si lega bene a monte della scatola TATA. Pertanto, è possibile sostituire 10 fattori di trascrizione con una singola proteina di fusione dCas9-AD e 10 sgRNA (RNA guida singola) per un punteggio di complessità totale di S = 11. È facile e veloce sintetizzare dieci sgRNA, mentre, come precedentemente commentato, l’assemblaggio di 10 proteine richiederebbe un lavoro molto più lungo e complicato.
In alternativa, si potrebbero usare due proteine dCas ortogonali (ad esempio, dCas9 e dCas12a): una per fondersi con un AD, e l’altra nuda o in combinazione con un dominio di repressione. Il punteggio di complessità aumenterebbe di una sola unità (S = 12). Quindi, i sistemi CRISPR-dCas sono la chiave per la costruzione di circuiti digitali genici molto complessi in S. cerevisiae.
Questo documento caratterizza profondamente l’efficienza dei repressori e degli attivatori basati su dCas9 e dCas12a nel lievito. I risultati mostrano che non richiedono un’elevata quantità di sgRNA per ottimizzare la loro attività, quindi i plasmidi episomiali sono preferenzialmente evitati. Inoltre, gli attivatori basati su dCas9 sono molto più efficaci quando si utilizza uno scaffold RNA (scRNA) che recluta copie del VP64 AD. Al contrario, dCas12a funziona bene se fuso direttamente al potente VPR AD. Inoltre, un promotore sintetico attivato richiede un numero variabile di siti bersaglio, a seconda della configurazione dell’attivatore (ad esempio, tre quando si utilizza dCas12a-VPR, sei per dCas9-VP64 e solo uno con dCas9 e uno scRNA). Come repressore, dCas12a appare più incisivo quando si lega la regione codificante piuttosto che il promotore.
Come svantaggio, tuttavia, CRISPR-dCas9 / dCas12a non interagiscono direttamente con le sostanze chimiche. Pertanto, potrebbero non essere utili nel livello di input. Per questo motivo, sono stati studiati progetti alternativi di gate booleani contenenti proteine anti-CRISPR (Acrs). Gli Acr agiscono sulle proteine (d)Cas e ne inibiscono il funzionamento8. Quindi, sono un mezzo per modulare l’attività dei sistemi CRISPR-(d)Cas. Questo articolo analizza a fondo le interazioni tra il tipo II Acrs e (d)Cas9, così come il tipo V Acrs e (d)Cas12a in S. cerevisiae. Poiché gli Acr sono molto più piccoli delle proteine Cas, è stato costruito un gate NOT sensibile all’estrogeno β-estradiolo fondendo il dominio legante l’ormone del recettore degli estrogeni umani9-HBD (hER) ad AcrIIA4. Inoltre, sono state realizzate una manciata di porte SÌ e NON che esprimevano costitutivamente dCas12a(-AD) e AcrVA dopo induzione con galattosio. Al momento, queste porte servono solo come prova di concetto. Tuttavia, rappresentano anche il primo passo verso un profondo ripensamento dell’algoritmo per effettuare la progettazione automatica computazionale di circuiti digitali genici sintetici nelle cellule di lievito.
