2011年,研究人员提出了一种计算方法,并开发了相应的软件,用于数字合成基因电路的自动设计1。用户必须指定输入数量(三个或四个)并填写电路真值表;这提供了使用电子技术推导电路结构的所有必要信息。真值表通过 Karnaugh 映射方法2 转换为两个布尔公式。每个布尔公式都由描述电路输入(部分)之间的逻辑运算(和或乘法)及其否定(文字)的子句组成。子句依次相加(OR)或乘法(AND)以计算电路输出。每个电路都可以根据其两个相应的公式中的任何一个来实现:一个以POS(总和乘积)形式编写,另一个以SOP(产品总和)表示形式编写。前者由包含文字逻辑和的子句(即布尔门)的乘法组成。相比之下,后者是文字相乘的子句的总和。
电路可以在面包板上通过将不同的门物理连接在一起来实现。电流允许门之间交换信号,从而计算输出。
在生物学中,情况更为复杂。布尔门可以实现为转录单元(TU;即真核细胞内的序列“启动子编码区域终止子”),其中转录或翻译(或两者)受到调节。因此,至少有两种分子建立了生物线路:转录因子蛋白和非编码、反义RNA。
基因数字电路被组织成两层或三层门,即:1)输入层,由YES(缓冲器)和NOT门组成,并将输入化学物质转换为布线分子;2) 内层,它由与相应布尔公式中的子句一样多的 TU 组成。如果电路按照SOP公式设计,则内层中的每个子句都将在所谓的分布式输出架构中产生电路输出(例如荧光)。如果使用和积(POS)公式,则需要3)最后一层,它将包含一个从内层收集布线分子的乘法门。
总体而言,在合成生物学中,可以为同一电路设计许多不同的方案。它们在TU和连接分子的数量和种类上有所不同。为了选择在酵母细胞中实现的最简单解决方案,每个电路设计都与复杂性评分S相关联,定义为
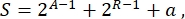
其中 A 代表激活剂的数量, R 代表阻遏剂的数量, a 是反义RNA分子的数量。如果电路中没有激活剂或抑制因子,则它们对S的贡献为零。因此,当需要大量正交转录因子时,在实验室中实现电路方案(高S)更加困难。这意味着新的活化器和抑制器应 重新 设计,以实现数字电路内部的完整布线。原则上,可以使用锌指蛋白3 和TAL效应子4 作为模板来组装新型DNA结合蛋白。但是,此选项似乎过于艰巨和耗时;因此,人们应该主要依靠小RNA和翻译调控来完成复杂的基因回路。
最初,这种方法是为了在细菌中制造数字电路而开发的。事实上,在真核细胞中,比起反义RNA,更适合谈论microRNA(miRNA)或小干扰RNA(siRNA)5。然而,RNAi途径不存在于 酿酒酵母中。因此,应该选择完全转录网络。假设一个电路需要五个激活器和五个抑制器;其复杂性分数为 S = 32。通过将10个转录因子替换为融合到激活域(AD)的单个dCas96 (核酸酶缺陷型Cas9),可以降低电路复杂性。如7所示,当在TATA盒和TSS(转录起始位点)之间结合启动子时,dCas9-AD在酵母中充当阻遏剂,当在TATA盒上游结合时,作为激活剂起作用。因此,可以用单个dCas9-AD融合蛋白和10个sgRNA(单个向导RNA)替换10个转录因子,总复杂性得分为S = 11。合成十种sgRNA既快速又容易,而如前所述,组装10种蛋白质需要更长、更复杂的工作。
或者,可以使用两种正交的dCas蛋白(例如dCas9和dCas12a):一种融合到AD,另一种裸露或与抑制结构域组合。复杂度分数只会增加一个单位 (S = 12)。因此,CRISPR-dCas系统是 酿酒酵母中构建非常复杂的基因数字电路的关键。
本文深入表征了酵母中基于dCas9和dCas12a的阻遏剂和激活剂的效率。结果表明,它们不需要大量的sgRNA来优化其活性,因此优先避免使用游离体质粒。此外,当使用募集VP64 AD拷贝的支架RNA(scRNA)时,基于dCas9的激活剂更有效。相比之下,dCas12a在直接融合到强VPR AD时效果很好。此外,根据激活剂的配置,合成活化启动子需要可变数量的靶位点(例如,使用 dCas12a-VPR 时为 3 个,dCas9-VP64 为 6 个,dCas9 和 scRNA 时只有一个)。作为阻遏因子,dCas12a在结合编码区而不是启动子时显得更敏锐。
然而,作为一个缺点,CRISPR-dCas9 / dCas12a不直接与化学物质相互作用。因此,它们在输入图层中可能没有用处。出于这个原因,已经研究了含有抗CRISPR蛋白(Acrs)的替代布尔门设计。Acrs作用于(d)Cas蛋白并抑制其工作8。因此,它们是调节CRISPR-(d)Cas系统活性的一种手段。本文深入分析了酿酒酵母II型Acrs与(d)Cas9以及V型Acrs和(d)Cas12a之间的相互作用。由于Acrs比Cas蛋白小得多,因此通过将人雌激素受体9-HBD(hER)的激素结合结构域融合到AcrIIA4,构建了对雌激素β-雌二醇有反应的NOT门。此外,在用半乳糖诱导时实现了少数表达dCas12a(-AD)的YES和NOT门和AcrVAs。目前,这些门仅用作概念证明。然而,它们也代表了深入重新思考算法的第一步,以执行酵母细胞中合成基因数字电路的计算自动设计。
