В 2011 году исследователи предложили вычислительный метод и разработали соответствующее программное обеспечение для автоматического проектирования цифровых синтетических генных схем1. Пользователь должен был указать количество входов (три или четыре) и заполнить таблицу истинности схемы; Это предоставило всю необходимую информацию для получения структуры схемы с использованием методов электроники. Таблица истинности была переведена в две логические формулы с помощью методакарты Карно 2. Каждая булева формула состоит из предложений, описывающих логические операции (сумма или умножение) между (частью) входов схемы и их отрицаний (литералов). Предложения, в свою очередь, либо суммируются (ИЛИ), либо умножаются (И) для вычисления выходного сигнала схемы. Каждая схема может быть реализована в соответствии с любой из двух соответствующих формул: одна записана в форме POS (произведение сумм), а другая – в представлении SOP (сумма произведений). Первый состоит из умножения предложений (т. е. булевых ворот), которые содержат логическую сумму литералов. Последний, напротив, представляет собой сумму предложений, в которых перемножаются литералы.
Электрические цепи могут быть реализованы на макетной плате путем физического соединения различных ворот вместе. Электрический ток позволяет обмениваться сигналами между вентилями, что приводит к вычислению выхода.
В биологии ситуация сложнее. Булевы ворота могут быть реализованы как транскрипционная единица (TU; т.е. последовательность «промотор-кодирующая область-терминатор» внутри эукариотических клеток), где транскрипция или трансляция (или и то, и другое) регулируются. Таким образом, по крайней мере, два вида молекул устанавливают биологическую связь: белки фактора транскрипции и некодирующие, антисмысловые РНК1.
Цифровая схема гена организована в два или три слоя вентилей, а именно: 1) входной слой, который состоит из затворов YES (буфер) и NOT и преобразует входные химические вещества в молекулы проводки; 2) внутренний слой, который состоит из стольких ТУ, сколько предложений в соответствующей булевой формуле. Если схема спроектирована в соответствии с формулой СОП, каждый пункт во внутреннем слое будет производить выходной сигнал схемы (например, флуоресценцию) в так называемой распределенной выходной архитектуре. Если используется формула произведения суммы (POS), то требуется 3) конечный слой, который будет содержать один мультипликативный затвор, собирающий молекулы проводки из внутреннего слоя.
В целом, в синтетической биологии для одной и той же схемы может быть разработано множество различных схем. Они различаются по количеству и виду как TU, так и молекул проводки. Чтобы выбрать самое простое решение, которое будет реализовано в дрожжевых клетках, каждая схема связана с оценкой сложности S, определяемой как
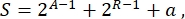
где A — количество активаторов, R — количество репрессоров, a — количество молекул антисмысловой РНК. Если в цепи отсутствуют активаторы или репрессоры, их вклад в S равен нулю. Следовательно, сложнее реализовать схему в лаборатории (высокий S), когда требуется большое количество ортогональных факторов транскрипции. Это означает, что новые активаторы и репрессоры должны быть спроектированы заново, чтобы реализовать полную проводку внутри цифровых цепей. В принципе, новые ДНК-связывающие белки могут быть собраны с использованием белков цинкового пальца3 и эффекторовTAL 4 в качестве шаблонов. Однако этот вариант представляется слишком трудным и трудоемким; поэтому следует полагаться в основном на малые РНК и регуляцию трансляции для завершения сложных генных цепей.
Первоначально этот метод был разработан для изготовления цифровых схем у бактерий. Действительно, в эукариотических клетках вместо антисмысловых РНК более уместно говорить о микроРНК (миРНК) или малых интерферирующих РНК (миРНК)5. Однако путь РНКи отсутствует у дрожжей S. cerevisiae. Следовательно, следует выбирать полностью транскрипционные сети. Предположим, что для цепи требуется пять активаторов и пять репрессоров; его оценка сложности будет S = 32. Сложность схемы может быть уменьшена путем замены 10 факторов транскрипции одним dCas96 (Cas9 с дефицитом нуклеазы), слитым с активационным доменом (AD). Как показано нарисунке 7, dCas9-AD работает как репрессор у дрожжей при связывании промотора между коробкой TATA и TSS (начальным сайтом транскрипции) и как активатор при связывании значительно выше по течению от коробки TATA. Таким образом, можно заменить 10 факторов транскрипции одним слитым белком dCas9-AD и 10 сгРНК (одиночными направляющими РНК) для общей оценки сложности S = 11. Синтезировать десять сгРНК можно быстро и легко, в то время как, как отмечалось ранее, сборка 10 белков потребовала бы гораздо более длительной и сложной работы.
В качестве альтернативы можно использовать два ортогональных белка dCas (например, dCas9 и dCas12a): один для слияния с AD, а другой голый или в сочетании с доменом репрессии. Оценка сложности увеличится всего на одну единицу (S = 12). Следовательно, системы CRISPR-dCas являются ключом к построению очень сложных цифровых схем генов у S. cerevisiae.
В данной работе подробно охарактеризована эффективность репрессоров и активаторов на основе dCas9 и dCas12a у дрожжей. Результаты показывают, что они не требуют большого количества сгРНК для оптимизации своей активности, поэтому эписомальные плазмиды предпочтительно избегают. Более того, активаторы на основе dCas9 гораздо более эффективны при использовании каркасной РНК (скРНК), которая рекрутирует копии VP64 AD. Напротив, dCas12a хорошо работает при непосредственном слиянии с сильным AD VPR. Кроме того, синтетический активированный промотор требует переменного числа целевых сайтов, в зависимости от конфигурации активатора (например, три при использовании dCas12a-VPR, шесть для dCas9-VP64 и только один с dCas9 и скРНК). В качестве репрессора dCas12a кажется более острым при связывании кодирующей области, а не промотора.
Однако недостатком является то, что CRISPR-dCas9/dCas12a не взаимодействуют с химическими веществами напрямую. Поэтому они могут быть бесполезны на входном уровне. По этой причине были исследованы альтернативные конструкции булевых затворов, содержащие белки анти-CRISPR (Acrs). Acrs действуют на (d)Cas белки и ингибируют их работу8. Следовательно, они являются средством модуляции активности систем CRISPR-(d)Cas. В данной работе подробно анализируются взаимодействия между типом II Acrs и (d)Cas9, а также типом V Acrs и (d)Cas12a у S. cerevisiae. Поскольку Acrs намного меньше, чем белки Cas, ворота NOT, реагирующие на эстроген β-эстрадиол, были построены путем слияния гормонсвязывающего домена рецептора эстрогеначеловека 9-HBD (hER) – с AcrIIA4. Кроме того, было реализовано несколько вентилей ДА и НЕ, которые конститутивно экспрессировали dCas12a(-AD) и AcrVA при индукции с галактозой. В настоящее время эти ворота служат только доказательством концепции. Тем не менее, они также представляют собой первый шаг к глубокому переосмыслению алгоритма для выполнения вычислительного автоматического проектирования цифровых схем синтетических генов в дрожжевых клетках.
