I 2011 foreslog forskere en beregningsmetode og udviklede et tilsvarende stykke software til automatisk design af digitale syntetiske genkredsløb1. En bruger skulle angive antallet af indgange (tre eller fire) og udfylde kredsløbets sandhedstabel; Dette gav alle de nødvendige oplysninger til at udlede kredsløbsstrukturen ved hjælp af teknikker fra elektronik. Sandhedstabellen blev oversat til to boolske formler via Karnaugh-kortets metode2. Hver boolsk formel er lavet af klausuler, der beskriver logiske operationer (sum eller multiplikation) blandt (en del af) kredsløbsindgange og deres negationer (de bogstavelige). Klausuler er igen enten opsummeret (OR) eller ganget (AND) for at beregne kredsløbsudgangen. Hvert kredsløb kan realiseres i henhold til en af dets to tilsvarende formler: den ene skrevet i POS (produkt af summer) form og den anden i SOP (sum af produkter) repræsentation. Førstnævnte består af en multiplikation af klausuler (dvs. boolske porte), der indeholder en logisk sum af de bogstavelige. Sidstnævnte er derimod en sum af klausuler, hvor de bogstavelige multipliceres.
Elektriske kredsløb kan realiseres på et brødbræt ved fysisk at forbinde forskellige porte sammen. Den elektriske strøm tillader udveksling af signaler mellem porte, hvilket fører til beregning af udgangen.
I biologi er situationen mere kompleks. En boolsk gate kan realiseres som en transkriptionsenhed (TU; dvs. sekvensen “promotorkodende regionterminator” inde i eukaryote celler), hvor transkription eller oversættelse (eller begge) reguleres. Således etablerer mindst to slags molekyler en biologisk ledning: transkriptionsfaktorproteinerne og de ikke-kodende, antisense RNA’er1.
Et gendigitalt kredsløb er organiseret i to eller tre lag porte, nemlig: 1) inputlaget, der er lavet af JA (buffer) og IKKE porte og omdanner inputkemikalierne til ledningsmolekyler; 2) det indre lag, der består af så mange TU’er, som der er klausuler i den tilsvarende boolske formel. Hvis kredsløbet er designet i henhold til SOP-formlen, vil hver klausul i det indre lag producere kredsløbsudgangen (f.eks. Fluorescens) i en såkaldt distribueret outputarkitektur. Hvis produktet af sum (POS) formel anvendes, kræves der et 3) sidste lag, som vil indeholde en enkelt multiplikativ port, der samler ledningsmolekylerne fra det indre lag.
Samlet set kan der i syntetisk biologi designes mange forskellige ordninger til det samme kredsløb. De adskiller sig i antallet og typen af både TU’er og ledningsmolekyler. For at vælge den nemmeste løsning, der skal implementeres i gærceller, er hvert kredsløbsdesign forbundet med en kompleksitetsscore S, defineret som
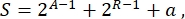
hvor A repræsenterer antallet af aktivatorer, R repræsenterer antallet af repressorer, og a er mængden af antisense RNA-molekyler. Hvis enten aktivatorer eller repressorer er fraværende fra kredsløbet, er deres bidrag til S nul. Derfor er det vanskeligere at realisere et kredsløbsskema i laboratoriet (højt S), når det kræver et stort antal ortogonale transkriptionsfaktorer. Dette betyder, at nye aktivatorer og repressorer skal konstrueres de novo for at realisere den komplette ledning inde i de digitale kredsløb. I princippet kan nye DNA-bindende proteiner samles ved hjælp af zinkfingerproteiner3 og TAL-effektorer4 som skabeloner. Denne mulighed virker imidlertid for besværlig og tidskrævende; derfor bør man hovedsagelig stole på små RNA’er og translationsregulering for at færdiggøre komplekse genkredsløb.
Oprindeligt blev denne metode udviklet til at fremstille digitale kredsløb i bakterier. Faktisk er det i eukaryote celler i stedet for antisense-RNA’er mere egnet at tale om mikroRNA’er (miRNA’er) eller små interfererende RNA’er (siRNA’er)5. RNAi-vejen er imidlertid ikke til stede i gæren S. cerevisiae. Derfor bør man vælge fuldt transkriptionsnetværk. Antag, at et kredsløb har brug for fem aktivatorer og fem undertrykkere; dens kompleksitetsscore ville være S = 32. Kredsløbskompleksiteten kan reduceres ved at erstatte de 10 transkriptionsfaktorer med en enkelt dCas96 (nukleasemangel Cas9) smeltet sammen til et aktiveringsdomæne (AD). Som vist i7 fungerer dCas9-AD som en repressor i gær, når der bindes en promotor mellem TATA-boksen og TSS (transkriptionsstartstedet) og som aktivator, når den binder godt opstrøms for TATA-boksen. Således kan man erstatte 10 transkriptionsfaktorer med et enkelt dCas9-AD fusionsprotein og 10 sgRNA’er (single guide RNA’er) for en total kompleksitetsscore på S = 11. Det er hurtigt og nemt at syntetisere ti sgRNA’er, mens samlingen af 10 proteiner som tidligere kommenteret ville kræve meget længere og mere kompliceret arbejde.
Alternativt kan man bruge to ortogonale dCas-proteiner (f.eks. dCas9 og dCas12a): den ene til at smelte sammen til en AD, og den anden bar eller i kombination med et undertrykkelsesdomæne. Kompleksitetsscoren ville kun stige med én enhed (S = 12). Derfor er CRISPR-dCas-systemer nøglen til konstruktionen af meget indviklede gendigitale kredsløb i S. cerevisiae.
Dette papir karakteriserer dybt effektiviteten af både dCas9- og dCas12a-baserede repressorer og aktivatorer i gær. Resultaterne viser, at de ikke kræver en høj mængde sgRNA for at optimere deres aktivitet, så episomale plasmider undgås fortrinsvis. Desuden er dCas9-baserede aktivatorer langt mere effektive, når man bruger et stillads-RNA (scRNA), der rekrutterer kopier af VP64 AD. I modsætning hertil fungerer dCas12a godt, når den smeltes direkte sammen med den stærke VPR AD. Desuden kræver en syntetisk aktiveret promotor et variabelt antal målsteder afhængigt af aktivatorens konfiguration (f.eks. tre ved brug af dCas12a-VPR, seks for dCas9-VP64 og kun en med dCas9 og et scRNA). Som repressor virker dCas12a mere skarp, når man binder kodningsregionen frem for promotoren.
Som en ulempe interagerer CRISPR-dCas9/dCas12a dog ikke direkte med kemikalier. Derfor er de muligvis ikke til nogen nytte i inputlaget. Af denne grund er alternative boolske gate-design, der indeholder anti-CRISPR-proteiner (Acrs), blevet undersøgt. Acrs virker på (d)Cas-proteiner og hæmmer deres arbejde8. Derfor er de et middel til at modulere aktiviteten af CRISPR-(d)Cas-systemer. Dette papir analyserer grundigt interaktionerne mellem type II Acrs og (d) Cas9, såvel som type V Acrs og (d) Cas12a i S. cerevisiae. Da Acrs er meget mindre end Cas-proteiner, blev en NOT-port, der reagerer på østrogen-β-østradiol, bygget ved at fusionere det hormonbindende domæne af den humane østrogenreceptor9-HBD (hER) til AcrIIA4. Desuden blev en håndfuld JA og IKKE porte, der udtrykte dCas12a (-AD) konstitutivt og AcrVA’er ved induktion med galactose, realiseret. På nuværende tidspunkt tjener disse porte kun som et bevis på konceptet. Men de repræsenterer også det første skridt mod en dyb nytænkning af algoritmen til at udføre det beregningsmæssige automatiske design af syntetiske gendigitale kredsløb i gærceller.
