気候変動と再生可能な資源の予測可能な枯渇は、私たちの社会に深刻な脅威をもたらします。この文脈において、酵素触媒は、持続可能で「環境に優しい」化学1の開発のため、まだ十分に活用されない可能性を表しています。酸化還元酵素は、穏やかな反応条件下で官能基を導入し、修飾を触媒し、最も重要な生体触媒2に所属する能力を有します。ほとんどの酸化還元変換は、NAD(P)Hのような補因子の外部の電源を必要とします。補因子再生のための方法は、工業規模で適用されています。しかし、彼らはまだほとんど価値の高い製品への適用を制限する、高い処理コストをもたらします。興味深いことに、いくつかのペルオキシダーゼ3,4およびP450モノオキシゲナーゼ5は 、いわゆる過酸化物のシャントを介して過酸化水素から電子を受け入れます。 H 2 O 2が安い共試薬であるが、それは伝えharmfあります多くの酵素のためのul。 その場の形成が着実に 過酸化水素の低濃度の酵素の安定動作を損なうことなく、反応を駆動するための実行可能なアプローチです。
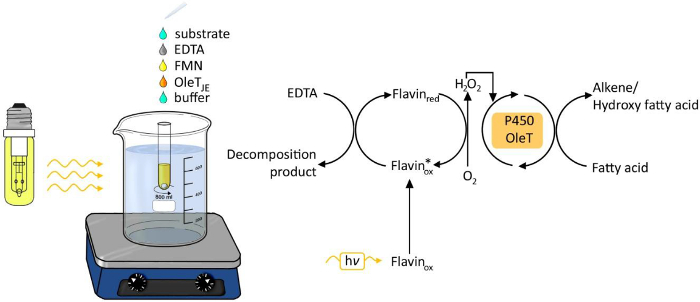
OLET JEによって脂肪酸のphotobiocatalytic脱炭酸の図1.実験のセットアップ。 この図の拡大版をご覧になるにはこちらをクリックしてください。
化学的および生物学的プロセスのためのエネルギー源として光を使用することは、最後の年6に増加注目されています。過酸化水素の光駆動世代レドックス変換( 図1)のために過酸化水素を供給することが容易かつ強固な方法として登場しました。このようなフラビンアデニン月として光触媒onucleotide(FMN)を酵素oxyfunctionalization反応の補因子として使用される過酸化水素に分子酸素の還元を可能にします。可能な電子供与体は、エチレンジアミン四酢酸(EDTA)、アスコルビン酸又は安価なギ酸です。この方法は、ペルオキシダーゼ3,4およびP450モノオキシゲナーゼ5を含むH 2 O 2依存性酵素のための一般的に適用可能です。
我々は最近、オレフィン8への天然脂肪の転換のための新規の細菌カルボキシラーゼ7の適用を検討しました。これは、バイオベースのソースから広く使用されているプラットフォームの化学物質の合成のための持続可能な経路であろう。グラム陽性菌Jeotgalicoccus SPからカルボキシラーゼOLET JE。脂肪酸の酸化的脱炭酸を触媒し、製品としての1-アルケンを形成します。 OLET JEは、細菌P450モノオキシゲナーゼに密接に関係していると、電子fは必要反応のためのロム過酸化水素。
残念ながら、基質と酵素の溶液にH 2 O 2の添加は、おそらくOLET JEの安定性に対する過酸化水素の有害な影響に、低い転化率及び結果の乏しい再現性をもたらしました。 NADPH還元RhFredとの融合タンパク質の生成が可能なNADPH依存性脱炭酸を行った。 図9は、それにもかかわらず、NADPHおよび費用効率的な再生のための現在の限られた可能性の高い価格が安く電子供与体を調査するために私たちを促しました。 P450モノオキシゲナーゼとOLET JEの類似性に触発され、我々は、H 2 O 2の光触媒による世代を使用していました。私たちは、無細胞抽出物または精製された酵素液を使用して(> 95%まで)、高い転化率を得るために喜んでいました。
脂肪酸脱炭酸の例では、我々は光駆動enzymのための一般的なプロトコルを提示します補因子として、光触媒と過酸化水素としてFMNを使用して、ATICレドックス変換。提示される方法は、Eの組換え細胞における酵素の産生を含みますコリ酵素の精製、1-アルケンの合成および反応生成物の分析のためのアプリケーション。
