Un essai biologique in vivo à haute résolution sur l’hydratation du pollen à grain unique pour Arabidopsis thaliana
Summary
Une méthode améliorée pour mesurer les profils d’hydratation du pollen chez Arabidopsis thaliana est décrite ici. La nouvelle méthode offre une résolution plus élevée, est non invasive et est hautement reproductible. Le protocole représente un nouvel outil pour une dissection plus fine des processus qui régulent les premiers stades de la pollinisation.
Abstract
La reproduction sexuée chez les plantes à fleurs nécessite une interaction initiale entre le grain de pollen et la surface stigmatique, où un dialogue moléculaire est établi entre les partenaires en interaction. Des études portant sur diverses espèces ont révélé qu’une série de points de contrôle moléculaires régulent l’interaction pollen-stigmate pour s’assurer que seul le pollen compatible, généralement intraspécifique, réussit à effectuer la fécondation. Chez les espèces qui possèdent un « stigmate sec », comme la plante modèle Arabidopsis thaliana, le premier point de contrôle de compatibilité prézygote post-pollinisation est l’établissement de l’hydratation du pollen.
Cette phase de pollinisation est étroitement réglementée, les signaux du grain de pollen suscitant la libération d’eau du stigmate, permettant ainsi l’hydratation du pollen. La capacité de mesurer et de suivre avec précision l’hydratation du pollen au fil du temps est essentielle à la conception d’expériences visant à comprendre la régulation de cette étape critique de la reproduction. Les protocoles publiés utilisent fréquemment des fleurs qui ont été excisées de la plante mère, maintenues sur des milieux liquides ou solides et pollinisées en vrac.
Cet article décrit un essai biologique de pollinisation in vivo non invasif qui permet un suivi minute par minute de l’hydratation des grains de pollen individuels d’A. thaliana à haute résolution. Le test est hautement reproductible, capable de détecter des variations très subtiles des profils d’hydratation du pollen, et convient donc à l’analyse des mutants qui affectent les voies régulant la pollinisation. Bien que le protocole soit plus long que ceux décrits pour les pollinisations en vrac, la précision et la reproductibilité qu’il fournit, ainsi que sa nature in vivo , le rendent idéal pour la dissection détaillée des phénotypes de pollinisation.
Introduction
La reproduction sexuée réussie chez les angiospermes repose généralement sur le transfert de grains de pollen intraspécifiques de l’anthère au stigmate, soit à l’intérieur des individus ou entre eux (c.-à-d. pollinisation). Ce transfert de grains de pollen à une fleur réceptive est généralement médié par des pollinisateurs ou des facteurs abiotiques; En tant que tel, cela entraîne également fréquemment le dépôt de pollen hétérospécifique dans des conditions naturelles. À quelques exceptions près, la progression de la pollinisation par le pollen hétérospécifique est désavantageuse sur le plan de l’évolution, réduisant la capacité de reproduction en raison de la perte d’opportunités d’accouplement, la plupart des descendants hybrides qui en résultent ne se développant pas correctement ou étant stériles1. Ainsi, des mécanismes ont évolué pour bloquer la pollinisation par des pollens hétérospécifiques « incompatibles» 2. La reconnaissance rapide du pollen compatible est donc sans doute le processus le plus important dans les premiers stades de la reproduction sexuée chez de nombreuses plantes à fleurs.
Dans la famille des Brassicaceae, où les stigmates sont de type « sec », une série de points de contrôle moléculaires agissent à plusieurs stades du processus de reproduction régulant la pollinisation, de sorte que seul le pollen compatible est efficace. L’hydratation du pollen est l’un des points de contrôle les plus importants (Figure 1), car le pollen qui ne parvient pas à s’hydrater ne peut pas progresser pour produire un tube pollinique et ensuite livrer des spermatozoïdes au gamétophyte femelle. Fréquemment, les grains incompatibles ne franchissent pas ce premier point de contrôle de pollinisation et n’ont donc pas accès à l’eau stigmatisante3. Chez les membres de la famille des Brassicaceae, la reconnaissance du pollen se produit rapidement, la compatibilité étant établie dans les minutes suivant la fixation du grain de pollen au pistil 4,5. Ces dernières années, beaucoup de progrès ont été réalisés et nous commençons maintenant à comprendre les mécanismes moléculaires qui régulent les principaux points de contrôle de la pollinisation.
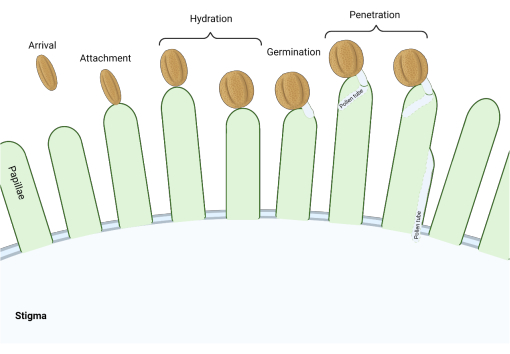
Figure 1 : Aperçu des événements clés lors de la pollinisation compatible. Ces étapes, telles que l’hydratation du pollen et la germination du tube pollinique, sont également des « points de contrôle » de pollinisation qui doivent être parcourus avec succès pour effectuer une pollinisation compatible. Le diagramme représente un stigmate de type « sec », typique des espèces de la familledes Brassicaceae 2,20. Veuillez cliquer ici pour voir une version agrandie de cette figure.
Des recherches pionnières sur le système d’auto-incompatibilité (SI) de Brassica, où le pollen « soi » est reconnu et rejeté, ont établi le paradigme de la reconnaissance de la stigmatisation pollinique chez les Brassicaceae 6,7,8,9,10. Le SI chez Brassica et ses parents est médié par des protéines de « reconnaissance » qui résident à la surface du pollen et à la membrane plasmique stigmatique qui, lors de l’interaction, conduisent au rejet du pollen. Le rejet du pollen SI opère par perturbation du système de compatibilité pollen-stigmate basal qui, lorsqu’il est pleinement activé par la perception du pollen compatible, conduit à une sécrétion ciblée par le stigmate, entraînant ainsi l’hydratation du pollen (pour les examens du mécanisme de compatibilité pollinique, voir11,12). Dans l’exemple du SI, le ligand transmis par le pollen est une petite protéine riche en cystéine, riche en cystéine S-locus (SCR/SP11), et le récepteur stigmatique est la kinase du récepteur S-locus (SRK).
Récemment, chez Arabidopsis thaliana, un autre groupe de petites protéines riches en cystéine transmises par le pollen, les protéines d’enveloppe pollinique de classe B (AtPCP-Bs), se sont révélées être des régulateurs importants de l’acceptation du pollen par l’activation de l’hydratation du pollen13. Les récepteurs stigmatiques des PCP-Bet les aspects de la voie réglementaire en aval ont également été décritsrécemment 14,15. Il est intéressant de noter que les études mutationnelles de gènes codant pour des médiateurs potentiels de signalisation du pollen transmis par le pollen et stigmatiques de l’hydratation du pollen (y compris AtPCP-Bs) n’ont pas réussi à générer des plantes qui ont un blocage complet au point de contrôle de l’hydratation du pollen. Cela suggère fortement que de multiples autres facteurs, encore non découverts, jouent un rôle dans la régulation de l’hydratation du pollen. En nous appuyant sur la méthode décrite pour la première fois par Wang et al.13, nous décrivons ici un essai biologique in vivo amélioré à haute résolution adapté à l’identification des défauts subtils d’hydratation du pollen dans les lignées candidates mutantes A. thaliana.
Protocol
Representative Results
Discussion
Pour les plantes à fleurs, les tout premiers stades de la reproduction sexuée sont sans doute les plus importants. Au niveau de l’interaction pollen-stigmate, des décisions moléculaires sont prises qui déterminent la « compatibilité » des partenaires en interaction. De telles décisions, si elles sont prises correctement, évitent le gaspillage de ressources qui pourrait avoir une incidence sur la capacité de reproduction21. Ainsi, permettre uniquement au pollen compatible d’effectuer la fécondation est un élément important du maintien de génotypes bien adaptés et, par conséquent, du succès évolutif des espèces. Les recherches menées avec la plante modèle A. thaliana ont été extrêmement précieuses pour approfondir notre compréhension de ce processus. Un certain nombre d’études au cours des dernières décennies ont révélé la présence de facteurs dans le manteau pollinique qui agissent au premier « point de contrôle » de compatibilité, où le pollen accède à de l’eau stigmatique pour permettre l’hydratation du pollen13. Malgré ces premières informations sur les mécanismes qui régulent la compatibilité pollen-stigmate, il existe encore de nombreuses lacunes dans notre compréhension de ce processus. À ce jour, aucun mutant de ligands transmis par le pollen ou de récepteurs stigmatiques connus pour avoir un impact sur l’hydratation du pollen ne peut bloquer complètement la pollinisation compatible, suggérant la présence d’autres déterminants non découverts de l’hydratation du pollen. En étant capable d’observer facilement le phénotype d’intérêt, le bioessai d’hydratation du pollen décrit ici est l’une des techniques les plus simples pour étudier les mutants potentiels qui régulent la pollinisation.
Les méthodologies existantes pour mesurer l’hydratation du pollen utilisent généralement des pollinisations en vrac et rapportent moins de points temporels 14,22,23, et peuvent donc manquer d’importants phénotypes de profil d’hydratation subtil. Par exemple, l’étude de Wang et al.13, ainsi que les travaux sur d’autres mutants de la protéine du manteau pollinique dans notre laboratoire (observations non publiées), ont révélé des différences intrigantes dans les profils d’hydratation entre les mutants. De telles différences subtiles peuvent contenir des indices importants sur les mécanismes de régulation sous-jacents à une pollinisation compatible.
La méthode décrite ici se concentre sur l’acquisition d’un nombre relativement faible de mesures entre les lignées végétales mutantes et WT, en mettant l’accent sur la précision méthodologique pour réduire la variation dans les ensembles de données. Bien que cette méthode soit hautement reproductible (comme le montre la figure 7), en supposant que la température et l’humidité sont correctement contrôlées, il est important de recueillir des données d’hydratation pour un nombre presque égal de pollen WT et mutant le même jour afin de réduire davantage le potentiel de variation. Les données peuvent ensuite être regroupées sur différents jours si nécessaire. En outre, la sélection des plantes de contrôle WT appropriées est essentielle pour une interprétation correcte des résultats d’hydratation. Pour le receveur de pollen, la même lignée végétale doit être utilisée pour recevoir à la fois des grains de contrôle WT et des grains de pollen mutants.
Par exemple, nous utilisons la lignée végétale mâle stérile pA9-barnase, qui est également présentée dans le protocole vidéo, comme destinataire du pollen WT (témoin) et mutant (expérimental) lors de l’étude des lignées mutantes de pollen d’ADN-T (telles que le mutant ‘KD’ décrit à la figure 8). Le mélange des données d’une telle lignée stérile masculine, qui n’a pas besoin d’être émasculée, avec celles recueillies à partir d’une lignée témoin émasculée manuellement doit être évité car ces stigmates se comporteront probablement différemment. De même, les lignées mutantes émasculées doivent être utilisées en conjonction avec une lignée WT émasculée (témoin) chaque fois que cela est possible. La même prudence doit également être appliquée lors de l’examen du patrimoine génétique des plantes étudiées. Alors que les collections les plus populaires de mutants d’ADN-T ont été générées dans le fond Col-0, d’autres, telles que la collection FLAG de l’Institut national de la recherche agronomique (INRA), sont disponibles dans le fond génétique de Wassilewskija (WS)24,25. Dans de tels cas, il est conseillé d’utiliser les lignées de plantes WT de l’écotype respectif comme témoins.
Bien que nous nous soyons concentrés ici sur l’hydratation du pollen au cours des 10 premières minutes de l’interaction pollen-stigmate, cette méthode peut également être adaptée pour englober des profils d’hydratation couvrant une période plus longue. Une caractéristique clé du protocole est que les fleurs restent attachées au courant végétal parent Les protocoles publiés nécessitent généralement l’excision du pistil et leur placement dans un milieu pour soutenir le tissu pendant la durée de l’expérience14,18,26. Bien qu’il n’y ait aucune preuve directe suggérant qu’une telle approche semi-in vivo ait un impact sur l’hydratation du pollen ou modifie même la régulation in vivo de ce processus, il est concevable que l’excision des fleurs de la plante mère puisse avoir un impact sur la pollinisation. Ainsi, ce protocole permet d’obtenir un véritable environnement in vivo pour l’étude de l’interaction pollen-stigmate, où l’intégrité structurelle de la plante est préservée.
Le transfert de grains de pollen uniques à des papilles stigmatiques « vierges » est sans doute l’une des opérations les plus difficiles décrites dans ce protocole. Il n’est pas rare de transférer des grappes de grains de pollen par erreur. Cependant, le risque que cela se produise peut être considérablement réduit en s’assurant que seule une monocouche de pollen est présente sur la pince (Figure 3A) (ou même un seul grain de pollen; Figure 5), et/ou en utilisant des grains de pollen déjà orientés, de sorte qu’ils « dépassent » des autres sur l’extrémité de la pince. Nous avons constaté qu’un opérateur expérimenté peut réussir le transfert d’un seul pollen vers une cellule papillaire stigmatique en environ 3 minutes et enregistrer des données pour jusqu’à cinq grains de pollen sur une période de 1 heure. Ainsi, sur une période de 2 à 4 jours, suffisamment de données peuvent être accumulées pour une analyse statistique significative des lignées végétales étudiées.
L’erreur humaine est potentiellement la plus grande source de variation dans l’analyse des ensembles de données dérivés d’études utilisant ce protocole. Par exemple, la définition de la « limite pollinique » lors de l’analyse de l’image dépend du jugement du chercheur individuel. Ainsi, il est possible que les mesures effectuées par différents chercheurs, même sur le même ensemble de données, puissent générer des variations. Dans la mesure du possible, un seul chercheur devrait effectuer les mesures afin de minimiser les erreurs d’échantillonnage. En outre, le couplage de l’analyse des ensembles de données WT et mutants par le même opérateur annule la définition potentiellement subjective de la « limite pollinique » et de la variation interopérateur.
En conclusion, une méthode sophistiquée mais précise pour mesurer les profils d’hydratation du pollen dans l’organisme modèle A. thaliana est décrite. Nous avons démontré qu’en utilisant ce protocole, des données très cohérentes sur l’hydratation du pollen pour A. thaliana peuvent être facilement acquises. Trois lots indépendants de données sur les pollinisations de WT acquises à des jours différents ont montré de petits écarts constants de <3% à tous les points temporels (figure 7 et tableau supplémentaire S1). Bien que l’essai biologique présenté ici soit légèrement plus complexe que la plupart des protocoles existants, la résolution des données générées est supérieure et convient à l’identification et à la caractérisation de nouveaux mutants qui ont un impact sur les voies régulant la pollinisation compatible.
Disclosures
The authors have nothing to disclose.
Acknowledgements
Cette recherche a été soutenue par des bourses d’études supérieures de l’Université de Bath (Université de Bath, Bath, Royaume-Uni, BA2 7AY) à Y.-L.L. et L.W. Figure 1 a été créé avec BioRender.com (https://biorender.com/).
Materials
| A9-barnase line | University of Bath | Courtsey of Prof. Rod Scott | Male sterile Arabidopsis thaliana wildtype equivalent line of the ecotype Columbia-0 |
| Dumont Tweezer, Dumont #5 Inox 11cm | Fisher | Dumont 500342 | Tweezer uses for transfer of pollen grain |
| GraphPad Prsim (version 8.0.2) | Dotmatics | Prism | Comprehensive data analysis, graphing and statistics software |
| JMP (version 17) | JMP Statistical Discovery LLC | JMP 17 | Statistical analysis software |
| Levington F2S seed & modular compost (with sand) | Levington | LEV75F2SMS | General-purpose compost for plant growth |
| Micromanipulator | Singer instrument Co. LTD. | Singer Micromanipulator | Micromanipulator to aid transfer of pollen grain |
| Nikon Digit sight DS-U1 | Nikon | DS-U1 | Microscope camera (coupletd to SMZ1500) |
| Nikon Eclipse TE2000-S Inverted Microscope | Nikon | TE2000-S | Inverted microscope |
| Nikon SMZ1500 Stereomicroscope | Nikon | SMZ1500 | Stereomicroscope |
| Nikon DS-Fi3 microscope camera | Nikon | DS-Fi3 | Microscope camera (coupletd to TE2000-S) |
| Nikon NIS-Elements Basic Research | Nikon | NIS-Elements BR | Image accquisition and analysis software (for DS-Fi3) |
| Nikon NIS-Elements F | Nikon | NIS-Elements F | Image accquisition and analysis software (for DS-U1) |
| WT Col-0 plant line | NASC | N700000 | Wildtype Arabidopsis thaliana, ecotype Columbia-0 |
References
- Rieseberg, L. H., Willis, J. H. Plant speciation. Science. 317 (5840), 910-914 (2007).
- Hiscock, S. J., Allen, A. M. Diverse cell signalling pathways regulate pollen-stigma interactions: the search for consensus. New Phytologist. 179 (2), 286-317 (2008).
- Kandasamy, M. K., Nasrallah, J. B., Nasrallah, M. E. Pollen pistil interactions and developmental regulation of pollen-tube growth in Arabidopsis. Development. 120 (12), 3405-3418 (1994).
- Bosch, M., Wang, L. Pollen-stigma interactions in Brassicaceae: complex communication events regulating pollen hydration. Journal of Experimental Botany. 71 (9), 2465-2468 (2020).
- Rozier, F., et al. Live-cell imaging of early events following pollen perception in self-incompatible Arabidopsis thaliana. Journal of Experimental Botany. 71 (9), 2513-2526 (2020).
- Dickinson, H. Dry stigmas, water and self-incompatibility in Brassica. Sexual Plant Reproduction. 8, 1-10 (1995).
- Takasaki, T., et al. The S receptor kinase determines self-incompatibility in Brassica stigma. Nature. 403 (6772), 913-916 (2000).
- Schopfer, C. R., Nasrallah, M. E., Nasrallah, J. B. The male determinant of self-incompatibility in Brassica. Science. 286 (5445), 1697-1700 (1999).
- Takayama, S., et al. Direct ligand-receptor complex interaction controls Brassica self-incompatibility. Nature. 413 (6855), 534-538 (2001).
- Shiba, H., et al. A pollen coat protein, SP11/SCR, determines the pollen S-specificity in the self-incompatibility of Brassica species. Plant Physiology. 125 (4), 2095-2103 (2001).
- Broz, A. K., Bedinger, P. A. Pollen-pistil interactions as reproductive barriers. Annual Review of Plant Biology. 72 (1), 615-639 (2021).
- Cheung, A. Y., Duan, Q., Li, C., James Liu, M. -. C., Wu, H. -. M. Pollen-pistil interactions: It takes two to tangle but a molecular cast of many to deliver. Current Opinion in Plant Biology. 69, 102279 (2022).
- Wang, L. D., et al. PCP-B class pollen coat proteins are key regulators of the hydration checkpoint in Arabidopsis thaliana pollen-stigma interactions. New Phytologist. 213 (2), 764-777 (2017).
- Liu, C., et al. Pollen PCP-B peptides unlock a stigma peptide-receptor kinase gating mechanism for pollination. Science. 372 (6538), 171-175 (2021).
- Bordeleau, S. J., Sanchez, L. E. C., Goring, D. R. Finding new Arabidopsis receptor kinases that regulate compatible pollen-pistil interactions. Frontiers in Plant Science. 13, 1022684 (2022).
- Suwabe, K., et al. Double-locking mechanism of self-compatibility in Arabidopsis thaliana: the synergistic effect of transcriptional depression and disruption of coding region in the male specificity gene. Frontiers in Plant Science. 11, 576140 (2020).
- Smyth, D. R., Bowman, J. L., Meyerowitz, E. M. Early flower development in Arabidopsis. Plant Cell. 2 (8), 755-767 (1990).
- Lee, H. K., Macgregor, S., Goring, D. R. A toolkit for teasing apart the early stages of pollen-stigma interactions in Arabidopsis thaliana. Pollen and Pollen Tube Biology. 2160, 13-28 (2020).
- Dilkes, B. P., et al. The maternally expressed WRKY transcription factor TTG2 controls lethality in interploidy crosses of Arabidopsis. PLoS Biology. 6 (12), 2707-2720 (2008).
- Riglet, L., et al. KATANIN-dependent mechanical properties of the stigmatic cell wall mediate the pollen tube path in Arabidopsis. eLife. 9, e57282 (2020).
- Zhou, L. Z., Dresselhaus, T. Friend or foe: Signaling mechanisms during double fertilization in flowering seed plants. Plant Development and Evolution. 131, 453-496 (2019).
- Gao, X. -. Q., et al. The Arabidopsis KINβγ subunit of the SnRK1 complex regulates pollen hydration on the stigma by mediating the level of reactive oxygen species in pollen. PLoS Genetics. 12 (7), e1006228 (2016).
- Lee, H. K., Goring, D. R. Two subgroups of receptor-like kinases promote early compatible pollen responses in the Arabidopsis thaliana pistil. Journal of Experimental Botany. 72 (4), 1198-1211 (2021).
- O’Malley, R. C., Barragan, C. C., Ecker, J. R. A user’s guide to the Arabidopsis T-DNA insertion mutant collections. Pollen and Pollen Tube Biology. 1284, 323-342 (2015).
- Samson, F., et al. FLAGdb++: a database for the functional analysis of the Arabidopsis genome. Nucleic Acids Research. 32, D347-D350 (2004).
- Doucet, J., et al. Investigations into a putative role for the novel BRASSIKIN pseudokinases in compatible pollen-stigma interactions in Arabidopsis thaliana. BMC Plant Biology. 19 (1), 549 (2019).
