Signaaltransductiewegen spelen een belangrijke rol in vrijwel elk celprocessen en de cel reageert snel te reageren op signalen uit de omgeving. 1 Deze trajecten worden vaak veroorzaakt door de binding van een signalerende molecule aan een extracellulaire receptor, hetgeen resulteert in activatie van intracellulaire enzymen. Amplificatie en vermeerdering van dit signaal in de cel wordt gemedieerd door de werking van signaaleiwitten die een netwerk van eiwit-eiwit interacties waarbij enzymen reversibel zijn geactiveerd met hoge specificiteit te vormen. Vanwege ontregeling van deze netwerken vaak leidt tot de ontwikkeling van kanker, is er veel belangstelling voor oprichting van 'signaaltransductie behandeling van kanker', 2 waarbij drugs zijn ontworpen om kwaadaardige signaalwegen verstoren geweest. We hebben onlangs voorgesteld een alternatieve benadering van transductie therapie die berust op het vermogen van geneesmiddelen om onnatuurlijke signaaltransductiewegen genereren signaal. <sup> 3 Met name zijn wij van mening dat door het ontwerpen van synthetische middelen die de functie van signaaleiwitten nabootsen, zou het mogelijk zijn om de functie van de cel indirect te moduleren. Bijvoorbeeld kunnen deze kunstmatige netwerken eiwitbiomarkers staat te splitsen enzymen die progeneesmiddelen activeren. Als alternatief kunnen deze signalering proteïne mimetica kunnen onnatuurlijke cel signaalwegen te activeren, wat resulteert in therapeutische effecten.
Om de uitvoerbaarheid van deze benadering tonen, hebben wij onlangs een synthetisch "chemisch transducer 4 dat bloedplaatjes afkomstige groeifactor (PDGF) maakt de splitsing van een kanker prodrug activeren door het activeren van glutathion-s-transferase (GST), wat niet zijn natuurlijke bindende partner. De structuur van deze 'transducer bestaat uit een anti-PDGF DNA aptameer dat is gemodificeerd met een tweewaardige remmer voor GST. Vandaar dit synthetische stof behoort tot een familie van moleculen met bindingsplaatsenverschillende eiwitten, 5-7 zoals chemische inductoren van dimerisatie (CID) 8-10 alsmede de zogenaamde eiwit-bindmiddel op basis van synthetische oligonucleotide-molecuul conjugaten. 11-21
De algemene beginselen van het ontwerpen van dergelijke systemen wordt hierin beschreven en gedetailleerde protocollen voor het synthetiseren en testen van de functie van dit 'transducer met gebruikelijke enzymatische assays worden voorzien. Dit werk ter vergemakkelijking ontwikkelen van aanvullende 'transducers' van deze klasse, die kunnen worden gebruikt om intracellulair eiwit-eiwit mediëren communicatie en dus aan kunstmatige cel signaalwegen induceren.
Figuur 1 beschrijft schematisch de werkingsprincipes van synthetische "chemische transducers 'dat onnatuurlijke eiwit-eiwit communicatie kan mediëren. In deze illustratie, een "chemische transducer, die synthetische bindmiddelen voor prot integreerteins I en II (bindmiddelen I en II), zodat het eiwit II katalytische activiteit van proteïne I, die niet de natuurlijke bindingspartner veroorzaken. Aangezien eiwit II, de omzetter verbindt de katalytische plaats van het enzym (eiwit I) en remt zijn activiteit (figuur 1, staat ii). De binding van de "transducer eiwit II echter bevorderen van de wisselwerkingen tussen bindmiddel I en het oppervlak van eiwitten II (figuur 1, staat iii) die zijn affiniteit richting eiwit I. de effectieve concentratie van minder Hierdoor de ' vrije 'transducer in de oplossing wordt gereduceerd, hetgeen leidt tot dissociatie van het transducer-eiwit I complex en reactivering van eiwit I (figuur 1, staat iv). Tezamen bieden deze stappen te markeren drie fundamentele uitgangspunten van het ontwerp van een efficiënte 'transducers': (1) een 'transducer' moet een specifieke binder hebben voor elk van de eiwit targets, (2) de interactie between bindmiddel II en eiwit II moet sterker dan de wisselwerking tussen bindmiddel I en I eiwit, en (3) binder I moet kunnen aangaan met het oppervlak van eiwitten II zijn. Dit laatste beginsel niet noodzakelijkerwijs dat bindmiddel ik alleen zou een hoge affiniteit en selectiviteit in de richting van eiwitten II hebben. In plaats daarvan is het gebaseerd op onze recente studies waaruit bleek dat het brengen van een synthetisch molecuul in de nabijheid van een eiwit waarschijnlijk interactie tussen deze moleculen en het oppervlak van het proteïne bevorderen. 19,22,23
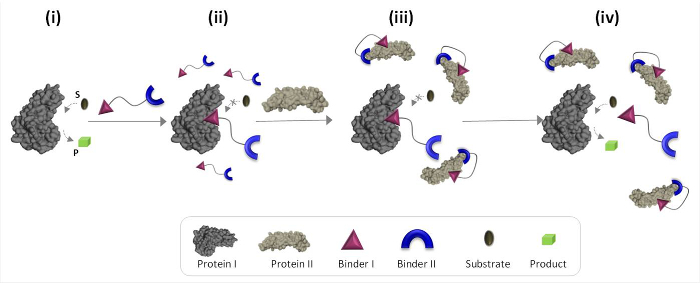
Figuur 1:. Werkingsprincipes van "chemische transducers 'Wanneer de" chemische transductor "toegevoegd actief eiwit I (toestand i), het bindt aan de actieve site via bindmiddel I en remt zijn activiteit (state ii). In de aanwezigheid van eiwit II echter de ongebonden 'chemische transducer "interageert met eiwit II tot bindmiddel II, die interacties tussen bindmiddel I en het oppervlak van eiwitten II bevordert. Dit veroorzaakte bindmiddel I-eiwit II interactie vermindert de effectieve concentratie van bindmiddel I, wat leidt tot dissociatie van het 'transducer'-eiwit I complex en eiwit I reactivering (state iv). Klik hier om een grotere versie van deze figuur te bekijken .
