Трансдукции сигнала пути играют важную роль практически во всех клеточных процессов и позволяют клетке быстро реагировать на внешние сигналы. 1 Эти пути часто вызываются связывание сигнальной молекулы с внеклеточным рецептором, что приводит к активации внутриклеточных ферментов. Усиление и распространение этого сигнала внутри клетки опосредовано функцией сигнальных белков, которые образуют сеть белок-белковых взаимодействий, в котором ферменты обратимо активированную с высокой специфичностью. Поскольку дисрегуляция этих сетей часто приводит к развитию рака, наблюдается большой интерес к созданию "сигнальной трансдукции терапии рака ', 2 ' в результате чего препараты предназначены для разрушения злокачественных сигнальных путей. Недавно мы предложили альтернативный подход к сигнальной трансдукции терапии, который зависит от способности препаратов генерировать неестественные пути передачи сигнала. <suр> 3 В частности, мы считаем , что при разработке синтетических агентов , которые имитируют функцию сигнальных белков, можно было бы модулировать функцию клетки косвенно. Например, эти искусственные сети могут позволить белка биомаркеров активировать ферменты, расщепляющие пролекарств. В качестве альтернативы, эти сигнальные белковые миметики могут быть способны активировать неестественные сигнальных путей клеток, в результате чего терапевтический эффект.
Для того, чтобы продемонстрировать возможность такого подхода, мы недавно создали синтетический 'химический датчик' 4 , который позволяет тромбоцитарный фактор роста (PDGF) , чтобы вызвать расщепление противоопухолевого пролекарства путем активации глутатион-S-трансферазы (GST), который не его естественный партнер по связыванию. Структура этого «датчика» состоит из анти-PDGF ДНК аптамеров, который модифицирован с ингибитором двухвалентной для GST. Следовательно, этот синтетический агент принадлежит к семейству молекул с сайтами связывания дляразличные белки, 5-7 , таких как химические индукторы димеризации (ИДС) 8-10 , а также к группе белков-связующие на основе олигонуклеотидных конъюгатов-синтетической молекулы. 11-21
Общие принципы, лежащие в основе конструкции таких систем описаны здесь и подробные протоколы для синтеза и тестирования функции этого «датчика» с обычными ферментативных анализов предоставляются. Эта работа предназначена для облегчения разработки дополнительных «преобразователей» этого класса, который может быть использован в качестве посредника внутриклеточный связи белок-белок, и, следовательно, для индукции искусственных сигнальных путей клеток.
На рисунке 1 схематически описывает принципы работы синтетических «химических преобразователей» , которые могут опосредовать неестественные белок-белковые связи. На этой иллюстрации, а 'химический преобразователь », который объединяет синтетические связующие вещества для PROTEins I и II (связующие вещества I и II), позволяет белок II, чтобы вызвать каталитическую активность белка I, который не является ее естественным партнером связывания. При отсутствии белка II, преобразователь связывает каталитический участок фермента (белка I) и ингибирует его активность (Рисунок 1, стенд II). Связывание "преобразователя" к белку II, тем не менее, обеспечивает взаимодействие между связующей I и поверхностью белка II (рис 1, стенд III), что снижает его сродство к белковой I. В результате эффективная концентрация ' свободный 'датчик в растворе уменьшается, что приводит к диссоциации преобразователя-белка I комплекса и реактивации белка I (рис 1, состояние IV). Взятые вместе, эти шаги выделить три основные принципы, лежащие в основе создания эффективных 'преобразователей': (1) 'датчик' должен иметь конкретное связующее для каждого из белковых мишеней, (2) взаимодействие betweен вяжущего II и белка II должен быть сильнее, чем взаимодействие между связующей I и белка I, и (3) вяжущего я должен иметь возможность взаимодействовать с поверхностью белка II. Этот последний принцип не обязательно требует, чтобы связующее только я бы высокое сродство и избирательность по отношению к белку II. Вместо этого, оно основано на наших недавних исследований , которые показали , что в результате чего синтетическую молекулу , в непосредственной близости к белку может способствовать взаимодействие между этой молекулой и поверхностью белка. 19,22,23
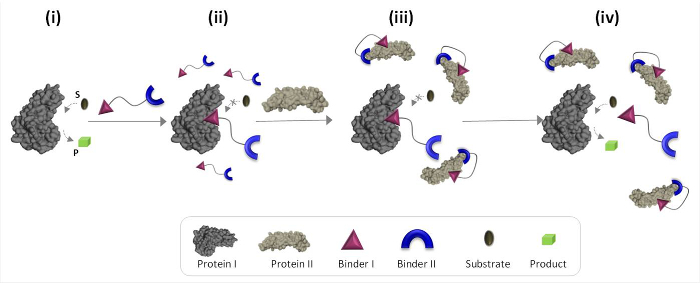
Рисунок 1:. Принцип работы технологии 'химических преобразователей "Когда" химический преобразователь' добавляется в активный белок I (состояние I), он связывается с его активным сайтом через связующее I и ингибирует его активность (состояние II). В присутствии белка II, однако, несвязанный "химический тransducer 'взаимодействует с белком II через вяжущего II, который обеспечивает взаимодействие между связующей I и поверхностью белка II. Это побудило связующее I-II белок взаимодействие снижает эффективную концентрацию связующего вещества I, что приводит к диссоциации "transducer'-белка I комплекса и белка I реактивации (состояние IV). Пожалуйста , нажмите здесь , чтобы посмотреть большую версию этой фигуры ,
