حيث ذكر إرنست [آب] في 1873، القرار للفحص المجهري الخفيفة التقليدية قد تم ويعتقد أن تقتصر على حوالي 200 شمال البحر الأبيض المتوسط بسبب حيود الضوء من الهدف1،2. حاليا تقنيات الفحص المجهري الخفيفة فائقة القرار كسر هذا القيد والسماح بالتقاط الصور الديناميكية مع القرار حيود الفرعية (< 200 nm). التقنيات عموما تندرج في فئتين عريضتين: حفز الانبعاثات استنفاد (STED) مجهرية على أساس النهج التي تولد حجم الإضاءة الحيود الفرعية بسبب الاستجابة البصرية غير الخطية من فلوروفوريس في3من العينات؛ ومن فوتواكتيفاتيد الخفيفة الميكروسكوب (النخيل) والتعمير البصرية العشوائية مجهرية (العاصفة)-على أساس تقنيات فائقة القرار، التي تستخدم الدوال الرياضية تعريب سينترويدس فلوروفوريس وثم إعادة تشكيل هذه سينترويدس لنموذج القرار فائقة الصور4،5. حاليا، بسبب الإعداد الضوئية غير معقدة نسبيا، النخيل والعاصفة على نطاق واسع يعملون بتفعيل فقط مجموعة فرعية صغيرة من فلوروفوريس في كل إطار من شريط فيديو طويل لتحضير البيولوجية. وهذا يسمح للترجمة أكثر دقة بملائمة غاوسي 2D من بقعة مضيئة، يسمى الدالة نقطة انتشار (PSF)، البروتينات المسماة فلوريسسينتلي في كل إطار من الفيديو. ثم يمكن تركيب موقع 2D كل جزيء المسمى فلوريسسينتلي على متن طائرة تصوير واحد لإنتاج صورة فائقة قرار لإعداد البيولوجية1،2. بينما هذه الترجمة جزيء واحد، نهج القرار فائقة للفحص المجهري ثورة التأكيد كيف أنجز تصوير العينات البيولوجية، ولا تزال هناك تحديات يجب التغلب عليها. على سبيل المثال، العاصفة والنخيل يمكن تحقيق أفضل الحلول المكانية الخاصة بها بعد تثبيت العينات البيولوجية وهكذا يقدم تمثيل ثابت للبروتينات المسمى فلوريسسينتلي، ووجود قيود مماثلة على الميكروسكوب الإلكتروني. بالإضافة إلى ذلك، تحقيق عالية الدقة المكانية لكل البروتين المسمى فلوريسسينتلي في الخلايا الحية، يجب تصويرها عينات في فراميراتس طويلة جداً وغير قادر على التقاط ديناميات البروتين. ولذلك، من الضروري للتغلب على هذه العقبات التقنية الرئيسية.
للحصول على دقة عالية الزمانية المكانية التي مناسبة تماما لكشف البروتينات أو الكشف تتحرك بسرعة في الخلايا الحية، وقد وضعنا القرار فائقة السرعة مجهرية في لدينا مختبر (الشكل 1)6،7، 8-عدة أوجه التقدم التقني الرئيسية في سرعة الفحص المجهري مكنتنا سابقا لتعقب النقل نوكليوسيتوبلاسميك من الجزيئات الصغيرة بنجاح، والبروتينات، ومرناً والفيروسات من خلال الأم النووية المسام المجمعات (الشخصيات)6، 7 , 8-بإيجاز، سيتم استخدام الميزات التالية لسرعة الفحص المجهري لتعقب الجزيئات تتحرك بسرعة من خلال الهياكل الفرعية ميكرومتر التناوب متناظرة في الخلايا الحية، مثل الشخصيات واهداب الأولية: (1) يميل أو الإضاءة الرأسية PSF تمكن إثارة جزيئات مفردة داخل وحدة تخزين صغيرة حيود حد في المستوى البؤري (الشكل 1)؛ (2) PSF يميل يمكن تجنب الأسفار خارج نطاق التركيز إلى حد كبير ومن ثم تحسين نسبة الإشارة إلى الضجيج. (3) كثافة بصرية من 100-500 كيلو واط/سم2 في الإضاءة PSF يسمح آلاف فوتونات التي سيتم جمعها من فلوروفوريس واحدة مع الكشف بسرعة بسرعة (> 500 هرتز). (4) الصيام الكشف عن سرعة يقلل إلى حد كبير أيضا خطأ التعريب المكانية جزيء مفرد (< 10 نانومتر) في تحديد مسارات المكانية تتحرك الجزيئات الفلورسنت في الخلايا الحية، نظراً للانتشار الجزيئي أحد العوامل الرئيسية تسبب عيوب التعريب جزيء واحد لنقل الجزيئات. (5) الراسخة في 2D إلى 3D التحويل خوارزميات تمكننا من توفير خرائط ثلاثية الأبعاد المكانية الكثافة الاحتمالية لطرق النقل للجزيئات في المجلس الوطني أو هدب الأولية. الجدير بالذكر أن لدينا عملية التحويل بين ديكارت ونظام التنسيق أسطواني يستخدم لتوليد كثافة الاحتمالية مكانية 3D خريطة بدلاً من 3D جزيء واحد تعقب (الشكل 2). سابقا، قد كشفت البيانات الميكروسكوب الإلكتروني أن9،المجلس الوطني10 و هدب الابتدائية11 كلا من بنية متناظرة التناوب. من حيث المبدأ، ينبغي نشرها الجزيئات تتحرك من خلال المجلس الوطني للصحافة أو هدب الأولية عشوائياً أيضا أن التوزيعات التناوب متناظرة. كما هو مبين في الشكل 2، عدد كبير من نشرها الجزيئات داخل الاسطوانة عشوائياً من شأنه أن يولد توزيعات التناوب متناظرة في عرض المقطع العرضي كالتي في المجلس الوطني، كما أدى نحو موحد المكانية التوزيع داخل كل منطقة فرعية صغيرة جداً بين اثنين من حلقات المجاورة (الشكل 2ه). ويؤدي هذا التوزيع الموحد أن التوزيع المكاني على طول البعد θ في نظام أسطواني ثابت. ثم يمكن تبسيط إحداثيات ثلاثي الأبعاد (R, X, θ) أن تكون إحداثيات 2D (R, X, ثابت). في الواقع، لدينا عملية التحويل بين نظامي أسطواني وديكارت من 2D (X, Y) 2D (R, X, ثابت). Θ ثابتة، تشير إلى الكثافة المكانية ف في الشكل 2ه، ويتم حسابها باستخدام المعادلة A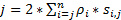 .
.
وفي نهاية المطاف، تتبع جزيء واحد له تطبيق واسع في البحوث البيولوجية، ومن ثم، فمن الطبيعي أن توضع مجموعة كبيرة تقنيات لسد منافذ البيولوجية المحددة12،،من1314. وهذا هو الحال بالنسبة لسرعة الفحص المجهري. سابقا، عندما يقترن مع خوارزمية تحول 3D، تم تطوير هذا الأسلوب لحل طرق النقل 3D لعبور الجزيئات من خلال اللجان التحضيرية الوطنية، هيكل البيولوجي التناوب متماثل والحجم diffraction الفرعية6. في هذه الورقة، تظهر أهداب الأولية لتكون نموذجا ممتازا العضيات كذلك. أهداب الأولية هي العضيات أسطواني، مثل هوائي (radius نانومتر ~ 125) أن المشروع من على سطح الأرض من الثدييات معظم الخلايا15،16،17. فمسؤولة عن تلقي إشارات خارجية، ويحيل بها استجابة داخل الخلايا المرتبطة عادة بالنمو والايض15،16. ولذلك، تدفق البروتينات الهيكلية، إعادة تدوير مستقبلات transmembrane، وانتقال الرسل داخل الخلايا المسؤوليات الحيوية من أهداب الأولية. في هذا المنعطف بين أهداب الابتدائي وجسم الخلية هو حاجز انتقائية حرجة، تسمى منطقة انتقالية أو TZ، من خلاله يجب أن يحدث كل هذا النقل البروتين11،،من1819، 20. بالإضافة إلى وظيفة TZ النابضة، واثنين على الأقل من عمليات النقل، النقل إينترافلاجيلار ونشر السلبي، يعتقد أن تكون مسؤولة عن حركة البروتين من خلال هذه المنطقة16،21، 22-من وجهة نظر صحة بشرية، فقدان أهداب الابتدائي وإلغاء الضوابط التنظيمية اللاحقة مما يشير إلى المصب مميزة لكثير من أنواع السرطان. وبالإضافة إلى ذلك، العديد من الأمراض الوراثية مثل متلازمة باردت-بيدل ومرض تكيس الكلي، ترتبط بالبروتين معيبة النقل23. جعل حجم الحد حيود الفرعية والعملية المعقدة للنقل البروتين انتقائية عن طريق TZ أهداب الابتدائي هدفا رئيسيا لهذه التقنية. في هذه الورقة، والأساليب وسوف نظهر تتبع البروتين transmembrane الهدبية، مستقبلات سوماتوستاتين 3 (SSTR3)24، المسمى خارجياً مع أليكسا فلور 647 ومكون من ايفت، IFT2025، المسمى مع جزيء تنصهر في التجارة والنقل.
