Da Ernst Abbe im Jahre 1873 erklärte, hat die Auflösung der konventionellen Lichtmikroskopie angenommen wurde, beschränkt sich auf etwa 200 nm durch Lichtbeugung aus dem Ziel1,2. Derzeit Höchstauflösung lichtmikroskopische Techniken brechen diese Einschränkung und ermöglicht die Erfassung von dynamischen Bildern mit Sub-Beugung (< 200 nm) Auflösung. Die Techniken in der Regel fallen in zwei Hauptkategorien: stimulierte Emission Depletion (STED) Mikroskopie-basierte Ansätze, die Sub-Beugung Beleuchtung Volumen durch nichtlineare optische Reaktion des Fluorophore Proben3; generieren und photoaktiviert Lichtmikroskopie (PALM) und stochastische optische Rekonstruktion Mikroskopie (STORM)-basierte Höchstauflösung Techniken, die mathematische Funktionen zum Zentroiden des Fluorophore lokalisieren und dann rekonstruieren diese Zentroide nutzen Super-Resolution Bilder4,5zu bilden. Derzeit sind aufgrund der relativ unkomplizierten optische Aufbau PALM und Sturm ausgiebig beschäftigt durch die Aktivierung nur einer kleinen Teilmenge der Fluorophore in jedem Frame ein langes Video eines biologischen Präparates. Dies ermöglicht genauere Lokalisierung von 2D “glockenförmig” Einbau von fluoreszierenden Ort bezeichnet die Point-spread-Funktion (PSF), Fluoreszent-markierten Proteine in jedem Frame des Videos. Die 2D Position jedes Molekül Eindringmittel beschriftet kann dann auf einen einzigen Abbildungsebene Höchstauflösung Bilder des biologischen Vorbereitung1,2überlagert werden. Während diese Lokalisierung Einzelmolekül-Höchstauflösung Ansätze zur Mikroskopie sicherlich revolutioniert wie Bildgebung von biologischen Proben durchgeführt wurde, gibt es noch Herausforderungen zu bewältigen. Beispielsweise können Sturm und PALM erreichen ihre beste räumlicher Auflösung nach der Fixierung von biologischen Proben und so präsentieren eine statische Darstellung der Fluoreszent-markierten Proteine, die eine ähnliche Einschränkung der Elektronenmikroskopie. Darüber hinaus müssen die Erreichung hohen räumlichen Auflösung für jedes Fluoreszent-markierten Protein in lebenden Zellen, Proben bei sehr langen Frameraten abgebildet werden die Proteindynamik erfassen können. Daher ist es notwendig, diese wichtigsten technische Hürden zu überwinden.
Um eine hohe räumlich-zeitliche Auflösung zu erhalten, die gut geeignet für die Erkennung von schnelllebigen Proteine oder RNA in lebenden Zellen ist, entwickelten wir Höchstauflösung Geschwindigkeit Mikroskopie in unserem Labor (Abbildung 1)6,7, 8. mehrere große technische Fortschritte in der Geschwindigkeit Mikroskopie haben zuvor konnten wir erfolgreich Nucleocytoplasmic Transport von kleinen Molekülen verfolgen, Proteine, mRNA und Virus durch Native nuclear pore komplexe (NPCs)6, 7 , 8. kurz, die folgenden Merkmale des SPEED-Mikroskopie werden zur schnelllebigen Makromoleküle durch Sub-Mikrometer rotationssymmetrisch Strukturen in lebenden Zellen, wie NPCs und primäre Zilien verfolgen: (1) eine geneigte oder eine vertikale Beleuchtung PSF ermöglicht die Anregung von einzelnen Molekülen in einem kleinen Beugungsgrenze Volumen in der Brennebene (Abbildung 1). (2), die geneigt PSF können stark vermeiden Sie Out-of-Focus Fluoreszenz und damit das Signal-Rausch-Verhältnis verbessern. (3) die optische Dichte von 100 bis 500 kW / cm2 bei der Beleuchtung PSF ermöglicht Tausende von Photonen aus einzelnen Fluorophore mit schnellen Erkennung Geschwindigkeiten (> 500 Hz) gesammelt werden. (4) die schnelle Erkennung Geschwindigkeit reduziert auch die räumliche Lokalisation Einzelmolekül-Fehler (< 10 nm) bei der Bestimmung der räumlichen Trajektorien fluoreszierende Moleküle in lebenden Zellen zu bewegen, weil molekulare Diffusion einer der wichtigsten Faktoren ist Unvollkommenheiten der Einzelmolekül-Lokalisierung für die Bewegung der Moleküle verursacht. (5) etablierte 2D zu 3D Umwandlung Algorithmen ermöglichen Ihnen 3D räumliche Wahrscheinlichkeitsdichte Karten der Transportwege für Moleküle in der NPC oder der primäre Zilie zu bieten. Es ist bemerkenswert, dass unsere Konvertierung zwischen den kartesischen und den zylindrischen Koordinatensystem verwendet wird, um ein 3D räumliche Wahrscheinlichkeitsdichte Karte anstatt 3D Einzelmolekül-Tracking (Abbildung 2) zu generieren. Bisher haben Elektronenmikroskopie Daten zeigten, dass die NPC9,10 und die primäre Zilie11 beide eine rotationssymmetrische Struktur haben. Grundsätzlich sollte nach dem Zufallsprinzip diffundierende Moleküle durch die NPC oder primäre Zilie rotationssymmetrische Distributionen auch haben. Wie in Abbildung 2dargestellt, eine hohe Anzahl von nach dem Zufallsprinzip diffundierende Moleküle im Inneren des Zylinders erzeugt rotationssymmetrische Verteilungen der Querschnitt durch das in der NPC, weiter führt eine etwa gleichmäßige räumliche Verteilung innerhalb jeder sehr kleinen Sub-Region zwischen zwei benachbarten Ringen (Abb. 2E). Diese gleichmäßige Verteilung führt, dass die räumliche Verteilung entlang θ Dimension im zylindrischen System konstant ist. Dann können die 3D Koordinaten (X, R, θ) vereinfacht werden, um die 2D Koordinaten (R, X, konstante) werden. Eigentlich ist unsere Konvertierung zwischen den kartesischen und den zylindrischen Systemen von 2D (X, Y) in 2D (R, X, konstante). Die konstante θ bezieht sich auf die räumliche Dichte p in Abbildung 2E, wird berechnet, indem die Gleichung A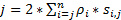 .
.
Letztlich Einzelmolekül-Tracking hat breite Anwendung in der biologischen Forschung, so ist es natürlich, dass eine Vielzahl von Techniken entwickelt, um spezifische biologische Nischen12,13,14füllen. Dies ist der Fall mit SPEED-Mikroskopie. Zuvor wurde zusammen mit einem 3D Transformationsalgorithmus diese Technik entwickelt, um 3D Transportwege von Transit-Moleküle durch die NPCs, eine sub-diffraction-Größe und rotationssymmetrische biologische Struktur6zu beheben. In diesem Papier primäre Zilien erweisen sich als ausgezeichnetes Modell Organellen sowie. Primäre Zilien sind zylindrisch, Antenne-wie Organellen (~ 125 nm Radius), die von der Oberfläche der meisten Säugetier-Zellen15,16,17zu projizieren. Sie sind verantwortlich für externe Signale empfangen und übertragen eine intrazelluläre Reaktion in der Regel verbunden mit Wachstum und Stoffwechsel15,16. Also flux von Strukturproteinen, recycling von transmembranen Rezeptoren und Übertragung von intrazellulären Botenstoffe sind wichtige Aufgaben der primären Zilien. An der Verbindungsstelle zwischen der primären Zilien und Zellkörper ist eine kritische Selektivität Schwelle, so genannte Übergangszone oder TZ, wodurch dieser Protein-Transport11,18,19erfolgen muss, 20. Neben der gating-Funktion von der TZ sind mindestens zwei Transportprozesse, intraflagellar Transport und passive Diffusion, vermutlich verantwortlich für die Bewegung des Proteins durch diese Region16,21, 22. aus Sicht der menschlichen Gesundheit, der Verlust der primäre Zilien und anschließenden Deregulierung der nachgeschalteten Signalisierung ist charakteristisch für viele Krebsarten. Darüber hinaus sind viele Erbkrankheiten wie Bardet-Bardet-Syndrom und polyzystischen Nierenerkrankung, defekte Protein Transport23zugeordnet. Die Größenbeschränkung von Sub-Beugung und den komplexen Prozess der selektiven Protein Transport durch die TZ machen die primären Zilien ein bevorzugtes Ziel für diese Technik. In diesem Methodenpapiers demonstrieren wir die Verfolgung von ciliary transmembranen Protein, Somatostatin Rezeptor 3 (SSTR3)24, extern mit Alexa Fluor 647 und Bestandteil des IFT, IFT2025, beschriftet mit einem geschmolzenen GFP-Molekül gekennzeichnet.
