Puisqu’a déclaré Ernst Abbe en 1873, la résolution de microscopie optique conventionnelle a cru être limitée à environ 200 nm en raison de la diffraction de la lumière de l’objectif1,2. Actuellement, les techniques de microscopie photonique Super-résolution briser cette limitation et permettent la capture d’images dynamiques avec résolution diffraction subsidiaire (< 200 nm). Les techniques généralement entrent dans deux grandes catégories : stimulée par des approches d’émission épuisement (STED) microscopie basée, qui génèrent des diffraction sous illumination volume en raison de la réponse optique non linéaire de fluorophores à échantillons3; et microscopie photonique photoactivation (PALM) et microscopie de reconstruction optique stochastique (tempête)-techniques de Super-résolution, qui utilisent des fonctions mathématiques pour localiser le centre de gravité des fluorophores et ensuite reconstituer ces centroïdes basées sur pour former la Super-résolution images4,5. Actuellement, en raison de la configuration optique relativement uncomplicated, PALM et tempête beaucoup travaillent en activant uniquement un petit sous-ensemble des fluorophores dans chaque image d’une vidéo longue d’une préparation biologique. Cela permet la localisation plus précise en 2D raccord gaussienne de la tache fluorescente, appelé la fonction de point propagation (PSF), de protéines marquées fluorescent dans chaque image de la vidéo. L’emplacement 2D de chaque molécule marquée fluorescent peut ensuite être superposé sur un seul plan d’imagerie pour produire une image de Super-résolution de la préparation biologique1,2. Tandis que ces molécules simples de localisation, Super-résolution approches à la microscopie certainement a révolutionné la mode d’exécution de l’imagerie d’échantillons biologiques, il y a encore des défis à surmonter. Par exemple, STORM et PALM peuvent réaliser leurs meilleures résolutions spatiales après fixation d’échantillons biologiques et ainsi présenter une représentation statique des protéines fluorescent marqué, qui est une restriction semblable de la microscopie électronique. En outre, pour parvenir à une résolution spatiale élevée pour chaque protéine fluorescent marqué dans des cellules vivantes, des échantillons doivent être copiés à des framerates très longs qui sont incapables de saisir la dynamique des protéines. Par conséquent, il est nécessaire pour surmonter ces obstacles techniques principales.
Pour obtenir une haute résolution spatio-temporelle qui est bien adaptée pour détecter les protéines rapides ou ARN dans des cellules vivantes, nous avons développé la microscopie de Super-résolution vitesse dans notre laboratoire (Figure 1)6,,7, 8. plusieurs avancées techniques majeures en microscopie de vitesse précédemment nous ont permis de suivre avec succès les transport nucléocytoplasmique de petites molécules, protéines, ADN messagère et virus par native nucléaire pore complexes (NPC)6, 7 , 8. en bref, les fonctionnalités suivantes de microscopie de vitesse seront utilisées pour suivre les macromolécules rapides par le biais de structures cône d’éclairage symétrique sous micromètre dans des cellules vivantes, telles que les PNJ et les cils primaires : (1) une tendance ou un l’éclairage vertical PSF permet l’excitation des molécules simples dans un volume de la petite limite de diffraction dans le plan focal (Figure 1) ; (2), les fibres discontinues de polyesters incliné peut éviter grandement fluorescence out-of-focus et donc d’améliorer le rapport signal-bruit. (3), la densité optique de 100 à 500 kW / cm2 dans l’illumination PSF permet à des milliers de photons à collecter des fluorophores unique avec des vitesses de détection rapide (> 500 Hz). Vitesse de détection (4), le jeûne réduit aussi considérablement l’erreur de localisation spatiale de molécules simples (< 10 nm) pour déterminer les trajectoires spatiales des molécules fluorescentes en mouvement dans des cellules vivantes, car la diffusion moléculaire est l’un des principaux facteurs imperfections de la localisation de la molécule unique pour le déplacement des molécules à l’origine. Algorithmes de transformation (5) bien établie 2D à la 3D pour nous permettent de fournir des cartes de densité de probabilité spatiale 3D des itinéraires de transport pour les molécules dans le NPC ou cil primaire. Il convient de noter que notre processus de conversion entre le cartésien et le système de coordination cylindrique est utilisé pour générer un suivi de molécules simples densité de probabilité spatiale 3D carte plutôt que 3D (Figure 2). Auparavant, la microscopie électronique données ont révélé que le NPC9,10 et le cil primaire11 tous les deux ont une structure par rotation symétrique. En principe, diffusant au hasard des molécules se déplaçant à travers le NPC ou cil primaire devrait également avoir des distributions de cône d’éclairage symétrique. Comme illustré à la Figure 2, un nombre élevé de diffusion au hasard des molécules à l’intérieur de la bouteille générerait des distributions cône d’éclairage symétrique à la vue de coupe transversale que celle dans le NPC, encore ayant pour résultat une sensiblement uniforme spatiale distribution au sein de chaque sous-région infime entre deux anneaux voisins (Figure 2E). Cette répartition uniforme entraîne que la distribution spatiale le long de la dimension θ dans le système cylindrique est constante. Puis les coordonnées 3D (R, X, θ) peuvent être simplifiées pour les coordonnées 2D (R, X, constante). En fait, notre processus de conversion entre le cartésien et les systèmes cylindriques est de la 2D (X, Y) en 2D (R, X, constante). La constante θ, fait référence à la densité spatiale p dans la Figure 2E, est calculée en utilisant l’équation A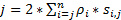 .
.
En fin de compte, suivi de la molécule unique a large application dans la recherche biologique, il est donc naturel qu’une pléthore de techniques est développée pour remplir les niches biologiques spécifiques12,13,14. Tel est le cas avec la microscopie de vitesse. Auparavant, lorsqu’il est couplé avec un algorithme de transformation 3D, cette technique a été développée pour résoudre les voies de transport 3D de molécules à travers les PNJ, une symétrie de révolution et sous-sous-diffraction-taille de la structure biologique6en transit. Dans cet article, cils primaires sont avérés organites excellent modèle aussi bien. Cils primaires sont des organites cylindriques, antenne-like (rayon de ~ 125 nm) qui projettent de la surface des cellules mammifères plus15,16,17. Ils sont responsables de recevoir des signaux externes et de transmettre une réponse intracellulaire, généralement associée à la croissance et le métabolisme15,16. Par conséquent, du flux des protéines structurales, recyclage des récepteurs transmembranaires et la transmission des messagers intracellulaires est des responsabilités vitales des cils primaires. À la jonction entre les cils primaires et le corps cellulaire est une barrière de sélectivité critique, appelée zone de transition ou TZ, grâce auquel tous les transports de cette protéine doivent avoir lieu à11,18,19, 20. En plus de la fonction blocage de la TZ, au moins deux processus de transport, intraflagellaire transport et diffusion passive, sont considérés comme responsables de la circulation des protéines à travers cette région16,,21, 22. du point de vue de la santé humaine, la perte des cils primaires et déréglementation ultérieure de la signalisation en aval est caractéristique de nombreux cancers. En outre, beaucoup de maladies génétiques, comme le syndrome de Bardet-Biedl et maladie polykystique des reins, est associés à la protéine défectueuse transport23. La taille limite de diffraction sous tant le processus complexe de transport des protéines sélectif par le biais de la TZ faire les cils primaires une cible de choix pour cette technique. Dans cet article des méthodes, nous allons démontrer le suivi d’une protéine transmembranaire ciliaire, récepteurs de la somatostatine 3 (SSTR3)24, marquées extérieurement à Alexa Fluor 647 ainsi qu’une composante de l’IFT, IFT2025, marquée avec une molécule GFP fusionnée.
