Como afirma Ernst Abbe en 1873, la resolución de la microscopía de luz convencional ha sido cree que limitada a aproximadamente 200 nm debido a la difracción de la luz del objetivo1,2. En la actualidad, técnicas de microscopía de superresolución rompen esta limitación y permiten la captura de imágenes dinámicas con la resolución de la secundario-difracción (< 200 nm). Las técnicas generalmente caen en dos amplias categorías: estimuló la emisión (STED) el agotamiento basado en la microscopia enfoques, generan volumen de iluminación sub-difracción debido a la respuesta óptica no lineal de fluoróforos en muestras3; y microscopia ligera fotoactivado (PALM) y microscopía óptica estocástica de la reconstrucción (tormenta)-basado en técnicas de súper-resolución, que utilizan funciones matemáticas para localizar los centroides de fluoróforos y luego reconstituir estos centroides para formar imágenes de súper-resolución4,5. Actualmente, debido a la configuración óptica relativamente sin complicaciones, Palma y tormenta son ampliamente empleados por activar sólo un pequeño subconjunto de fluoróforos en cada fotograma de un video largo de una preparación biológica. Esto permite la localización más exacta por ajuste gaussiano 2D del spot fluorescente, denominada la función de extensión de punto (PSF), de proteínas marcada con fluorescencia en cada fotograma del vídeo. La ubicación 2D de cada molécula marcada con fluorescencia puede entonces se superponen en un solo plano de proyección de imagen para producir una imagen de súper-resolución de la preparación biológica1,2. Mientras estos localización de una sola molécula, enfoques de super-resolución microscopía revolucionaron sin duda cómo se realizó la proyección de imagen de las muestras biológicas, todavía hay retos que superar. Por ejemplo, tormenta y la palma pueden alcanzar sus mejores resoluciones espaciales después de la fijación de muestras biológicas y así presentar una representación estática de las proteínas marcada con fluorescencia, que es una limitación similar de la microscopia electrónica. Además, para conseguir alta resolución espacial para cada proteína marcada con fluorescencia en células vivas, las muestras deben ser reflejadas en tasa de fotogramas muy largo que es capaces de captar la dinámica de la proteína. Por lo tanto, es necesario superar estos obstáculos técnicos principales.
Para obtener una alta resolución espacio-temporal que está bien preparado para detectar movimiento rápido proteínas o ARN en células vivas, hemos desarrollado la microscopía de superresolución velocidad en nuestro laboratorio (figura 1)6,7, 8. varios avances técnicos importantes en microscopía de velocidad previamente nos han permitido rastrear con éxito transporte NUCLEOCITOPLASMÁTICO de pequeñas moléculas, proteínas, mRNA y virus a través de nativos nuclear poro complejos (PNJ)6, 7 , 8. brevemente, se utilizarán las siguientes características de la microscopia de la velocidad para seguir el rápido movimiento de macromoléculas a través de estructuras rotationally simétrico secundario-micrómetro en células vivas, como NPCs y cilios primarios: (1) una inclinación o un iluminación vertical PSF permite la excitación de las moléculas individuales dentro de un volumen pequeño del límite de difracción en el plano focal (figura 1); (2) el PSF inclinado puede evitar grandemente la fluorescencia fuera de enfoque y así mejorar la relación señal a ruido. (3) la densidad óptica de 100-500 kW / cm2 en la iluminación PSF permite miles de fotones a recogerse en solo fluoróforos con velocidades de detección rápida (> 500 Hz). Velocidad de la detección (4) el ayuno también reduce el error de localización espacial de una sola molécula (< 10 nm) en la determinación de las trayectorias espaciales de mover moléculas fluorescentes en células vivas, ya que la difusión molecular es uno de los principales factores que causan las imperfecciones de la localización de una sola molécula de moléculas en movimiento. Algoritmos de transformación (5) consolidada 2D a 3D nos permiten ofrecer mapas de densidad de probabilidad espacial 3D de rutas de transporte para las moléculas en el NPC o el cilio primario. Cabe mencionar que nuestro proceso de conversión entre el cartesiano y el sistema de coordinación cilíndrico se usa para generar una densidad de probabilidad espacial 3D mapa en lugar de 3D sola molécula seguimiento (figura 2). Anteriormente, datos de microscopía electrónica han revelado que el NPC9,10 y el cilio primario11 ambos tienen una estructura de simetría de rotación. En principio, difusión al azar de moléculas en movimiento a través del NPC o cilio primario debe tener distribuciones rotationally simétricas. Como se muestra en la figura 2, un número elevado de difusión al azar de las moléculas dentro del cilindro podría generar distribuciones rotationally simétricas en la vista de corte transversal que en el NPC, aún más dando como resultado un aproximadamente uniforme espacial distribución dentro de cada subregión muy pequeño entre dos anillos vecinos (figura 2E). Esta distribución uniforme conduce que la distribución espacial a lo largo de la dimensión θ en el sistema cilíndrico es constante. Entonces las coordenadas 3D (X, R, θ) se pueden simplificar para las coordenadas 2D (R, X, constante). En realidad, nuestro proceso de conversión entre el cartesiano y los sistemas cilíndricos es de 2D (X, Y) 2D (R, X, constante). El θ constante, se refiere a la densidad espacial p en la figura 2E, se calcula utilizando la ecuación A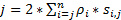 .
.
En definitiva, una sola molécula seguimiento tiene amplia aplicación en la investigación biológica, por lo tanto, es natural que se desarrollará una gran cantidad de técnicas para llenar nichos biológicos específicos12,13,14. Tal es el caso de microscopía de la velocidad. Anteriormente, cuando se combina con un algoritmo de transformación 3D, esta técnica se desarrolló para resolver rutas de transporte 3D de moléculas a través de los PNJs, una estructura biológica rotationally simétricos y sub-diffraction-tamaño6en tránsito. En este papel, cilios primarios aparecen ser organelos excelente modelo así. Cilios primarios son orgánulos cilíndricos, antena-como (~ 125 radio de nm) que se proyectan desde la superficie de las células mamíferas más15,16,17. Son responsables de recibir las señales externas y transmitir una respuesta intracelular típicamente asociada con el crecimiento y metabolismo15,16. Por lo tanto, del flujo de proteínas estructurales, reciclaje de receptores transmembranales y la transmisión de los mensajeros intracelulares son responsabilidades vitales de los cilios primarios. En la coyuntura entre los cilios primarios y el cuerpo celular es una barrera crítica de la selectividad, llamada zona de transición o TZ, a través del cual este transporte de la proteína debe ocurrir11,18,19, 20. Además de la función bloquea de la TZ, al menos dos procesos de transporte, Transporte intraflagelar y difusión pasiva, se cree que son responsables del movimiento de la proteína a través de esta región16,21, 22. desde un punto de vista de salud humana, la pérdida de cilios primarios y posterior desregulación de señalización corriente abajo es característica de muchos tipos de cáncer. Además, muchas enfermedades genéticas, como síndrome de Bardet-Biedl y enfermedad de riñón policística, están asociadas con la proteína defectuosa transporte23. El tamaño límite de sub- difracción y el complejo proceso de transporte de la proteína selectiva a través de la TZ hacen los cilios primarios el primer objetivo de esta técnica. En este trabajo métodos, demostramos el seguimiento de una proteína transmembrana ciliar, receptor de somatostatina 3 (SSTR3)24, marcado externamente con un componente de IFT, IFT2025, marcado con una molécula GFP fusionada y Alexa Fluor 647.
