Ernst Abbe tarafından 1873 yılında belirtilen beri konvansiyonel ışık mikroskobu çözünürlüğe objektif1,2ışık kırınım nedeniyle sınırlı yaklaşık 200 nm olduğuna inanılan. Şu anda, süper çözümleme ışık mikroskobu teknikleri bu sınırlama kırmak ve alt kırınım (< 200 nm) çözünürlük ile dinamik görüntü yakalama sağlar. Teknikleri, genellikle iki geniş kategoriye ayrılır: uyarılmış alt kırınım Aydınlatma Cilt fluorophores doğrusal olmayan optik tepki nedeniyle örnekleri3‘ te; oluşturmak emisyon tükenmesi (STED) mikroskopi dayalı yaklaşımlar ve photoactivated ışık mikroskobu (PALM) ve Stokastik optik imar mikroskobu (fırtına)-fluorophores cisimlerin yerelleştirilmesine ve bu cisimlerin yeniden oluşturmak için Matematiksel işlevler kullanmak süper çözümleme teknikleri dayalı süper çözünürlük görüntüleri4,5oluşturmak için. Şu anda, nispeten basit optik Kur nedeniyle PALM ve fırtına geniş yalnızca küçük bir alt kümesine fluorophores biyolojik hazırlanması uzun bir videonun her çerçevede etkinleştirerek istihdam edilmektedir. Bu daha doğru yerelleştirme için floresan spot, 2D Gauss montaj tarafından noktaya yayılmış işlevi (PSF) olarak adlandırdığı video her çerçevede fluorescently etiketli proteinlerin sağlar. Her fluorescently etiketli molekül 2D konumunu sonra biyolojik hazırlık1,2süper çözünürlük görüntü üretmek için tek bir görüntüleme uçağa bindirilen. Bu tek molekül yerelleştirme sırasında mikroskobu süper çözünürlük yaklaşımlar kesinlikle nasıl görüntüleme biyolojik örneklerin gerçekleştirilen devrim, hala aşılması gereken sorunlar vardır. Örneğin, fırtına ve PALM onların en iyi uzamsal çözünürlük sonra biyolojik örneklerin fiksasyonu elde etmek ve böylece fluorescently etiketli proteinler, statik bir temsili olan elektron mikroskobu benzer bir kısıtlamasıdır mevcut. Ayrıca, canlı hücrelerdeki her fluorescently etiketli protein için yüksek Uzaysal çözünürlük elde etmek için örnekleri protein dynamics yakalamak mümkün değildir çok uzun framerate yansıması gerekir. Bu nedenle, bu ana teknik engellerin üstesinden gelmek gereklidir.
Çok hızlı hareket eden protein veya RNA’ların canlı hücrelerde algılamak için uygundur yüksek bir kronolojik zamanmekansal çözünürlük elde etmek için biz süper çözünürlük hızı mikroskobu bizim laboratuvar (şekil 1)6,7‘, gelişen 8. hız mikroskobu birkaç önemli teknik gelişmeler bizi küçük moleküllerin nucleocytoplasmic taşıma başarılı bir şekilde izlemek daha önce etkinleştirdiyseniz, proteinler, mRNA ve virüs-den geçerek yerli nükleer gözenek kompleksleri (NPCs)6, 7 , 8. kısaca, hız mikroskobu, aşağıdaki özellikleri hızlı hareket eden oluştururlar alt mikrometre rotasyonel simetrik yapıları NPCs ve birincil kirpikler gibi canlı hücrelerdeki üzerinden izlemek için kullanılan: (1) bir eğimli veya bir Dikey aydınlatma PSF odak düzlemi (şekil 1); küçük kırınım-limit hacmindeki içinde tek moleküllerin uyarma sağlar (2) eğimli PSF büyük ölçüde odak floresan önlemek ve böylece sinyal-gürültü oranı geliştirmek. (3) 100-500 kW optik yoğunluk / cm2 aydınlatma PSF’hızlı algılama hızı (> 500 Hz) ile tek fluorophores üzerinden tahsil edilecek fotonlar binlerce sağlar. (4) hızlı algılama hızı da büyük ölçüde azaltır tek molekül kayma yerelleştirme hata (< 10 nm) moleküler Difüzyon önemli faktörlerden biri olduğu için canlı hücrelerde, floresan molekülleri hareket kayma yörüngeleri belirlemede molekülleri hareket etmek için tek molekül yerelleştirme kusurları neden. (5) well-established 2D 3D dönüştürme algoritmaları molekülleri NPC veya birincil cilium 3D mekansal olasılık yoğunluk haritalar ulaşım yollarının sağlamamıza olanak sağlar. Bu bizim dönüştürme işlemi Kartezyen ve silindirik koordinasyon sistemi arasında bir 3D mekansal olasılık yoğunluk 3D yerine harita tek molekül izleme (Şekil 2) oluşturmak için kullanılan dikkat çekicidir. Daha önce elektron mikroskobu veri-si olmak kâşif NPC9,10 ve11 birincil cilium hem de rotasyonel simetrik bir yapıya sahip. Prensip olarak, rasgele NPC veya birincil cilium aracılığıyla hareketli moleküllerin Difüzyon rotasyonel simetrik dağıtımları da olmalıdır. Şekil 2‘ de gösterildiği gibi rastgele silindir içinde moleküllerin Difüzyon yüksek bir dizi bu kesit görünümü, rotasyonel simetrik dağıtımları daha fazla bir yaklaşık Tekdüzen kaynaklanan NPC oluşturmak kayma Dağıtım iki komşu yüzük (Şekil 2E) arasında çok küçük her alt bölge içerisinde. Bu tekdüze dağılım silindirik sistemdeki θ boyuttaki kayma dağıtım sabittir yol açar. Sonra 3D koordinatları (R, X, θ)-ebilmek var olmak kolaylaştırmak-2B koordinatları (R, X, Sabit) olmak. Aslında, bizim dönüştürme işlemi Kartezyen ve silindirik sistemleri arasındaki 2D (R, X, Sabit) 2D (X, Y) etmektir. Sürekli θ, Şekil 2Emekansal yoğunluk p başvurduğu, ADenklem kullanılarak hesaplanır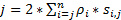 .
.
Sonuçta, tek molekül izleme geniş uygulama Biyolojik araştırma vardır, böylece, bu teknikleri bir bolluk belirli biyolojik nişler12,13,14doldurmak için geliştirilecek doğaldır. Böyle hız mikroskobu ile durumdur. Daha önce bir 3B dönüştürme algoritması ile birleştiğinde, moleküller NPCs, alt diffraction ölçekli ve rotasyonel simetrik biyolojik yapısı6üzerinden transit, 3D ulaşım yolları çözmek için bu teknik geliştirilmiştir. Bu yazıda, birincil kirpikler mükemmel model organelleri de olduğu gösterilmiştir. Birincil kirpikler en memeli hücreleri15,16,17yüzeyinden Project’in silindirik, anten gibi organellerin (~ 125 nm RADIUS) vardır. Onlar dış sinyalleri alma ve genellikle büyüme ve metabolizma15,16ile ilgili hücre içi bir yanıt gönderme için sorumludur. Bu nedenle, yapısal proteinlerin akı, geri dönüşüm transmembran reseptörler ve hücre içi haberci iletimini birincil kirpikler önemli sorumlulukları vardır. Dönemde birincil kirpikler ve hücre beden arasındaki geçiş bölgesi veya TZ, hangi aracılığıyla bu protein taşıma11,18,19ortaya çıkması gereken bir kritik seçicilik engel vardır, 20. TZ gating işleve ek olarak, en az iki taşıma işlemi, intraflagellar taşıma ve pasif difüzyon, protein bu bölge16,21, üzerinden hareket sorumlu olduğu düşünülür 22. bir insan sağlığı açısından birincil kirpikler kaybı ve aşağı akım sinyal sonraki deregülasyon birçok kanser özelliğidir. Buna ek olarak, Bardet-Biedl sendromu ve Polikistik böbrek hastalığı, gibi birçok genetik hastalıkların arızalı protein taşıma23ile ilişkilidir. Alt kırınım boyutu sınırı ve seçici protein taşıma TZ aracılığıyla karmaşık süreci birincil kirpikler için bu tekniği ilk hedef olun. Bu yöntemleri yazıda, biz silier transmembran protein, somatostatin reseptörü 3 (SSTR3) izleme gösterecektir24dışarıdan Alexa Fluor 647 ve bir bileşeni olan bir yuvarlak GFP molekül ile etiketli LFT, IFT2025, taşır,.
