以来、1873 年にエルンスト ・ アッベが述べたように、従来の光学顕微鏡の分解能は、客観的1,2から光の回折による限られた約 200 nm と考えられています。現在、超解像光学顕微鏡技術は、この制限を破るし、サブ回折 (< 200 nm) 解像度とダイナミックな画像を取得できます。技術は一般に 2 つの広範なカテゴリに分類: サンプル3で fluorophores の非線形光学応答のため、サブの回折照明ボリュームを生成、排出枯渇 (STED) 顕微鏡ベース アプローチを刺激センサーの光学顕微鏡観察 (パーム) と確率論的光再建顕微鏡 (嵐)-ベースの fluorophores の重心をローカライズし、この重心を再構成する数学的な関数を利用した超解像技術超解像画像4,5を形成。現在、比較的単純な光のセットアップのためパームと嵐が広く採用のみ生物学的製剤の長いビデオの各フレームでの同時の小さなサブセットをアクティブにします。これによりより正確な局在化のため 2次元ガウシアンフィッティング蛍光スポットの点拡がり関数 (PSF) と呼ばれる、ビデオの各フレームで蛍光標識したタンパク質の。蛍光標識した分子の 2次元位置は、生物学的準備1,2の超解像画像を生成する単一のイメージング平面上、重ね合わせることが。これらの分子の局在化しながら超解像顕微鏡方法確かに革命をどのように生体試料のイメージングを行った、まだ克服すべき課題があります。たとえば、嵐とパーム生体試料の固定後最高の分解能を達成できしたがって電子顕微鏡のような制限である蛍光標識したタンパク質の静的な表現を提示します。さらに、タンパク質のダイナミクスを捉えることである非常に長いフレーム レートで、生きた細胞の蛍光標識蛋白質ごとの高空間分解能を達成するためにサンプルをイメージする必要があります。したがって、これらの主要な技術的なハードルを克服するために必要です。
生細胞における高速移動蛋白質または Rna の検出に適しています高い時空間分解能を得るためには、私たち研究室 (図 1)6,7,の超解像顕微鏡を開発しました。8. 顕微鏡のいくつかの主要な技術の進歩は正常に小分子の核細胞質輸送を追跡する私たちを有効にしていた、ネイティブ核を介してウイルス mRNA、タンパク質孔複合体 (Npc)6,7,8. Npc など一次繊毛の生細胞におけるサブミクロン回転対称形構造の高分子の高速移動を追跡する簡単に、顕微鏡の次の機能が使用されます: (1) 傾斜または垂直照明 PSF により焦点面 (図 1); 小さな回折限界ボリューム内単一分子の励起(2) 傾斜の PSF は大きくアウト フォーカス蛍光を回避でき、信号対雑音比を向上させる。100-500 kW の光学濃度 (3)/cm2照明 PSF の高速検出速度 (> 500 Hz) で単一 fluorophores が付いてから収集する光子の数千をことができます。(4) 高速検出速度も大幅分子拡散は 1 つの主要な要因の生細胞における蛍光分子の移動の空間軌道を決定する単一分子空間定位エラー (< 10 nm) が減少します。分子の移動の単一分子の局在化の欠陥を引き起こします。(5) 老舗 2D 3d 変換アルゴリズムは、NPC または一次繊毛中分子の輸送ルートの 3 D 空間確率密度マップを提供するために私たちを有効にします。それはデカルトと円筒の調整システムの間の変換処理を使用して、3 D 空間の確率密度 3 D ではなくマップ単一分子追跡 (図 2) を生成することは注目に値する。以前は、電子顕微鏡データでは NPC9,10と一次繊毛11両方回転対称形の構造があることを明らかにしました。原則として、ランダムに拡散分子 NPC または一次繊毛を移動する必要があります回転対称形の分布。図 2に示すように、ランダムにシリンダーの中の分子の拡散の数が多いがおよそ均一の結果さらに NPC の回転対称形分布と断面生成空間2 つの隣接リング (図 2E) 間の非常に小さい各サブ領域内で配布します。この均一な分布を導く円筒における θ 次元空間分布は一定であります。その後、3 D 座標 (R, X, θ) は 2 D 座標 (R, X, 定数) であると簡略化できます。実際には、デカルトと円筒形のシステムの間の変換プロセスは (X, Y) は 2 D から 2 D (R, X, 定数) です。一定の θ図 2電子の空間密度pを指し、 Aの方程式を使用して計算されます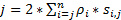 。
。
最終的には、単一分子追跡研究生物学の広範なアプリケーションには、したがって、特定の生物学的ニッチ12,13,14を入力するテクニックの茄多が開発することは当然です。顕微鏡の場合と同様であります。以前は、3 D 変換アルゴリズムと相まって、この手法が通過する Npc、diffraction サイズと回転対称の生物学的構造6による分子の 3 D 輸送ルートを解決する開発されました。本稿で一次繊毛がエクセレント モデル細胞小器官と同様に表示されます。一次繊毛は、プロジェクトのほとんどの哺乳類細胞15,16,17の表面からアンテナのように、円筒の器官 (~ 125 nm radius) です。外部信号を受信し、成長と代謝15,16に通常関連付けられている細胞内の応答を送信に責任があります。したがって、構造タンパク質のフラックス、膜貫通受容体と細胞内メッセンジャーの伝送のリサイクル一次繊毛の重要な責任です。一次繊毛、細胞体の間の連接は転移の地帯または11,18,19を発生する必要がありますこのすべてのタンパク質輸送、TZ と呼ばれる重要な選択性バリア 20。TZ のゲーティング機能に加えて少なくとも 2 つの輸送プロセス, 鞭毛内輸送と受動拡散この地域16,21,蛋白質の動きを担当すると考えられています。22. ヒトの健康の観点から主な毛損失と下流シグナルの後の規制緩和は多くの癌の特性。さらに、バルデ-ビードル症候群や多嚢胞性腎疾患など、多くの遺伝病は、欠陥のある蛋白質輸送23に関連付けられます。サブの回折限界サイズと TZ のたんぱく質の選択的輸送の複雑なプロセスの両方一次繊毛この技術のための主なターゲットを作る。この方法で毛様体膜貫通タンパク質、ソマトスタチン受容体 3 (SSTR3) の追跡を紹介24、Alexa Fluor 647 と IFT、IFT2025、溶かされた GFP の分子とラベルのコンポーネントで外部ラベルします。
