Понимание клеточных мембран и их функций замечательно изменилось за последние несколько десятилетий. Прежний взгляд на клеточные мембраны как на пассивные липидные бислои, которые определяют границы клеток и белки домашней мембраны1, постепенно трансформировался в динамическую модель, в которой липидные бислои играют важную роль в регулировании жизненно важных биологических процессов, включая клеточную сигнализацию, молекулярный обмен и функцию белка — и это2,3,4,5,6. Это осознание того, что клеточные мембраны очень динамичны, постоянно подвергаются ремоделированию и молекулярному перераспределению, побудило к научным исследованиям за пределами равновесных структур мембран7,8,9. Соответственно, было разработано несколько подходов к изучению различных динамических режимов в биологических и биоинспирируемых липидных мембранах. На сегодняшний день большинство этих исследований в основном сосредоточено на диффузных молекулярных движениях10,11,12,13 и макроскопических флуктуациях формы14,15,16,оставляя значительный пробел в понимании динамики промежуточных мембран, т. е. коллективных флуктуаций липидных сборок, состоящих из нескольких 10-100 молекул липидов. Эта динамика происходит на масштабах длины от нескольких десятков до нескольких 100 Å и во временных масштабах от суб-ns до нескольких сотен ns (см. Рисунок 1),называемых здесь мезоскопическими масштабами. Именно в этих масштабах происходит ключевая биологическая активность на мембранном уровне17. Это включает вирусное почкование18,канал19и мембранно-белковые взаимодействия20. Важно также отметить, что энергетический ландшафт мембранных белков21, 22показывает, что конформационные изменения в белках, необходимые для их регуляторной роли, происходят на временных масштабахns 23 коллективных флуктуаций мембран, что дополнительно подчеркивает важность мезоскопической динамики в биологической функции клеточных мембран и их биоинспирируемых аналогов20. В данной работе основное внимание уделяется двум основным мезоскопическим динамическим режимам в липидных мембранах, а именно флуктуациям изгиба и колебаниям толщины.
Основной проблемой при непосредственном провожде этих флуктуацийных режимов является сложность одновременного доступа к их пространственным и временным масштабам с использованием стандартных методов спектроскопии. Другая проблема заключается в том, что методы прямого контакта могут влиять на те же колебания, которые они предназначены для измерения16. Это еще более усугубляется композиционной и структурной сложностью биологических мембран24,25,что приводит к неоднородным особенностям мембраны, включая образование липидных доменов26,27, 28,29, 30и асимметрию мембран31, 32, 33,требуя селективных зондов для понимания динамики различных особенностей мембраны. К счастью, эти проблемы могут быть преодолены с помощью неинвазивных методов нейтронной спектроскопии, таких как нейтронное спиновое эхо (NSE), которые по своей сути получают доступ к требуемой длине и временным масштабам, а также позволяют дополнительно проводить исследования селективных особенностей мембран без изменения их физико-химической среды34. Действительно, за последние несколько лет спектроскопия NSE превратилась в уникальный и мощный зонд коллективной мембранной динамики35. Результаты исследований NSE на липидных мембранах дали новое представление о механическихсвойствах 36,37 и вязкоупругих38,39 липидных мембран и пролили новый свет на их потенциальную роль в биологической функции40,41.
Метод спектроскопии NSE основан на конструкции интерферометрического прибора, впервые предложенной Mezei42,с использованием серии спин-ласт и магнитных катушек для управления прецессией спина нейтрона, когда нейтроны пересекают инструмент. Конструкция опирается на магнитное зеркальное отражение элементов магнитного поля относительно положения образца(рисунок 1А). Это означает, что при отсутствии обмена энергией между нейтроном и образцом нейтрон выполняет одинаковое количество спиновых прецессий в противоположных направлениях в первой и второй половине инструмента (обратите внимание на π-флиппер между двумя катушками прецессии). В результате конечное спиновое состояние нейтрона остается неизменным относительно начального состояния – явление, называемое спин-эхо (см. прозрачный нейтрон на рисунке 1А). Однако, когда нейтрон энергетически взаимодействует с образцом, энергообмен изменяет число спиновых прецессий во второй половине инструмента, что приводит к другому конечному спиновому состоянию (см. Рисунок 1А). Это экспериментально обнаруживается как потеря поляризации, как будет показано далее в этой статье. Для получения более подробной информации о технике NSE читатель обращается к специальным техническим документам42,43,44,45.
Здесь представлено упрощенное описание, чтобы дать приблизительную оценку длины и временных масштабов, доступных с NSE. Масштабы длин определяются диапазоном достижимых волновекторных передач, Q = 4π sin θ/λ, где 2θ — угол рассеяния, а λ — длина волны нейтрона. Видно, что Q задается диапазоном длин волн и степенью вращения второго плеча спектрометра (см. рисунок 1А). Типичный Q-диапазонна спектрометрах NSE составляет ~0,02-2 Å-146,47и до 0,01-4 Å-1 с недавними обновлениями48,49,что соответствует пространственным масштабам ~1-600 Å. С другой стороны, доступная шкала времени вычисляется из общего угла прецессии (или фазы), приобретенного нейтроном в магнитных катушках прецессии, и обнаруживается, что она составляет50: 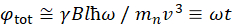 . В этом выражении t — время Фурье, определяемое как
. В этом выражении t — время Фурье, определяемое как 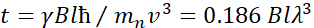 , где —
, где — 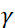 нейтронное гиромагнитное
нейтронное гиромагнитное 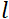 отношение, — длина катушки и
отношение, — длина катушки и 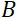 — сила магнитного поля катушки. Стоит отметить, что время Фурье — это величина, которая строго зависит от геометрии прибора, силы магнитного поля и длины волны нейтрона. Например, используя нейтроны длины волны
— сила магнитного поля катушки. Стоит отметить, что время Фурье — это величина, которая строго зависит от геометрии прибора, силы магнитного поля и длины волны нейтрона. Например, используя нейтроны длины волны 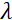 = 8 Å и настройки прибора = 1,2 м и = 0,4 Тл, время Фурье вычисляется как t ~ 50 нс. Экспериментально время Фурье настраивается путем изменения тока в катушках прецессии (т. Е. Напряженность магнитного поля) или с использованием различных длин волн нейтронов, что приводит к типичным шкалам времени NSE от ~ 1 пс до 100 нс. Тем не менее, недавние обновления спектрометров NSE позволили получить доступ к более длинным временам Фурье, до ~ 400 нс на спектрометре J-NSE-Phoenix в Центре Хайнца Майера-Лейбница51 и спектрометре SNS-NSE в Национальной лаборатории Оук-Ридж48и до ~ 1000 нс на спектрометре IN15 NSE в Институте Лауэ-Ланжевена (ILL)49.
= 8 Å и настройки прибора = 1,2 м и = 0,4 Тл, время Фурье вычисляется как t ~ 50 нс. Экспериментально время Фурье настраивается путем изменения тока в катушках прецессии (т. Е. Напряженность магнитного поля) или с использованием различных длин волн нейтронов, что приводит к типичным шкалам времени NSE от ~ 1 пс до 100 нс. Тем не менее, недавние обновления спектрометров NSE позволили получить доступ к более длинным временам Фурье, до ~ 400 нс на спектрометре J-NSE-Phoenix в Центре Хайнца Майера-Лейбница51 и спектрометре SNS-NSE в Национальной лаборатории Оук-Ридж48и до ~ 1000 нс на спектрометре IN15 NSE в Институте Лауэ-Ланжевена (ILL)49.
Помимо прямого доступа к длине и временной шкале мембранной динамики, NSE обладает присущими 52 возможностями чувствительности к изотопам нейтронов52. В частности, способность нейтронов по-разному взаимодействовать с изотопами водорода, наиболее распространенного элемента в биологических системах, приводит к различной плотности рассеяния нейтронов,34 или NSLD (эквивалент оптического показателя преломления50),когда протий замещается дейтерием. Это позволяет использовать подход, известный как вариация контраста, который обычно используется для выделения конкретных особенностей мембраны или сокрытия других — последний сценарий называется контрастным соответствием. Частым применением изменения/согласования контраста является замена воды (NSLD = -0,56 × 10-6 Å-2)тяжелой водой или D2O (NSLD = 6,4 × 10-6 Å-2)для усиления нейтронного сигнала от протиированных липидных мембран (NSLD ~ 0 × 10-6 Å-2). Этот подход очень эффективен в исследованиях структуры мембраны, поскольку проникновениеD2O в область головной группы мембраны позволяет точно определить толщину мембраны (см. Рисунок 2A,левая панель) и расположение различных липидных подгрупп при применении более сложных моделей53,54. В данной работе приводятся некоторые примеры использования вариации контраста для изучения коллективной динамики в биомиметических мембранах и отдельных особенностей мембран.
Здесь эффективность NSE в предоставлении уникального понимания динамических и функциональных свойств мембран иллюстрируется наглядными примерами исследований NSE на модельных и биологически значимых липидных мембранных системах с акцентом на мезомасштабную динамику в отдельно стоящих мембранах в виде липосомальных суспензий. Для NSE измерений динамики плоских мембран читатель обращается к специализированным публикациям по нейтронно-эхо-спектроскопии (GINSES)55, 56 и другим исследованиям выровненных многоламелярных мембранных стеков57,58,59,60.
Для простоты в данной работе выделены три различные схемы мембранной дейтрации, проиллюстрированные на хорошо изученной доменно-формирующей, или фазоразделяющей, липидной бислойной системе из смесей 1,2-димиристоил-sn-глицеро-3-фосфохолин (DMPC) и 1,2-дистеароил-sn-глицеро-3-фосфохолин (DSPC)61,62. Эти два липида характеризуются несоответствием длины углеводородной цепи (14 атомов углерода на хвост в DMPC против 18 атомов углерода на хвост в DSPC) и температуры перехода гель-жидкость (Tm, DMPC = 23 °C против Tm, DSPC = 55 °C). Это приводит к боковому фазовой разделению в мембранах DMPC:DSPC при температурах между верхними и нижними температурами перехода смеси63. Рассмотренные здесь схемы дейтерации выбраны для демонстрации различных динамических режимов, доступных в измерениях NSE на липосомальных мембранах, а именно флуктуаций изгиба, колебаний толщины и селективных флуктуаций изгиба/толщины боковых доменов. Все липидные композиции представлены для двухслоев DMPC:DSPC, приготовленных при моль фракции 70:30, с использованием коммерчески доступных протиированных и пердевтерированных вариантов DMPC и DSPC. Все этапы подготовки образца основаны на 4 мл липосомальной суспензии вD2Oс концентрацией липидов 50 мг/мл для общей липидной массы Mtot = 200 мг на образец.
