A compreensão das membranas celulares e sua função evoluiu notavelmente nas últimas décadas. A visão anterior das membranas celulares como bicamadas lipídicas passivas que definem limites celulares e proteínas de membranadoméstica 1 gradualmente se transformou em um modelo dinâmico no qual bicamadas lipídicas desempenham um papel importante na regulação de processos biológicos vitais, incluindo sinalização celular, troca molecular e função proteica — para citar alguns2,3,4,5,6. Essa constatação de que as membranas celulares são altamente dinâmicas, constantemente submetidas à remodelação e redistribuição molecular, tem incitado explorações científicas além das estruturas de equilíbrio das membranas7,8,9. Assim, várias abordagens foram desenvolvidas para estudar os diversos modos dinâmicos em membranas lipídicas biológicas e bioinspiradas. Até o momento, a maioria desses estudos se concentrou principalmente nos movimentos moleculares difusivos10,11,12,13 e flutuações de forma macroscópica14,15,16, deixando uma lacuna significativa na compreensão da dinâmica da membrana intermediária, ou seja, flutuações coletivas de conjuntos lipídicos consistindo de poucos 10-100s de moléculas lipídicas. Essas dinâmicas ocorrem ao longo de escalas de comprimento de poucas dezenas a poucos 100 Å e ao longo do tempo escalas de sub-ns a algumas centenas de ns (ver Figura 1), referidas aqui como escalas mesoscópicas. É nestas escalas que a atividade biológica chave ocorre no nível de membrana17. Isso inclui o brotamento viral18, gating canal19e interações membrana-proteína20. Também é importante ressaltar que a paisagem energética das proteínas da membrana21,22 mostra que as alterações conformais nas proteínas — necessárias para seu papel regulatório — acontecem ao longo das escalas de tempons 23 de flutuações coletivas de membrana, enfatizando ainda mais a importância da dinâmica mesoscópica na função biológica das membranas celulares e seus analógicos bioinspirados20. Este artigo se concentra nos dois modos dinâmicos mesoscópicos primários em membranas lipídicas, ou seja, flutuações de dobra e flutuações de espessura.
O principal desafio em sondar diretamente esses modos de flutuação é a dificuldade em acessar simultaneamente suas escalas espaciais e temporais usando métodos de espectroscopia padrão. O outro desafio é que as técnicas de contato direto podem impactar as mesmas flutuações que se destinam a medir16. Isso é ainda mais exacerbado pela complexidade composicional e estrutural das membranas biológicas24,25, o que resulta em características de membrana não homogênea, incluindo a formação de domínio lipídudo26,27,28,29,30 e assimetria de membrana 31,32,33– exigindo sondas seletivas para entender a dinâmica de diferentes características da membrana. Felizmente, esses desafios podem ser superados com métodos não invasivos de espectroscopia de nêutrons, como o echo de giro de nêutrons (NSE), que acessam inerentemente o comprimento e as escalas de tempo necessárias, e permitem ainda estudos de características de membrana seletiva sem alterar seu ambiente físico-químico34. De fato, nos últimos anos a espectroscopia NSE evoluiu para uma sonda única e poderosa da dinâmica da membrana coletiva35. Os resultados de estudos da NSE sobre membranas lipídicas produziram novos insights sobre as propriedades mecânica36,37 e viscoelástica38,39 de membranas lipídicas e lançaram uma nova luz sobre seu potencial papel na função biológica40,41.
A técnica de espectroscopia NSE é baseada em um design de instrumento interferométrico, proposto pela primeira vez por Mezei42, usando uma série de giradoras e bobinas magnéticas para controlar a precessão do giro de nêutrons à medida que os nêutrons atravessam o instrumento. O desenho repousa no espelhamento magnético dos elementos do campo magnético em relação à posição da amostra(Figura 1A). Isso implica que, na ausência de troca de energia entre o nêutron e a amostra, o nêutron realiza o mesmo número de precessões de spin, em direções opostas, na primeira e segunda metade do instrumento (observe o π-flipper entre as duas bobinas de pré-cessão). Como resultado, o estado final de rotação do nêutron permanece inalterado em relação ao estado inicial – um fenômeno referido como spin-echo (ver nêutron transparente na Figura 1A). No entanto, quando o nêutron interage energeticamente com a amostra, a troca de energia modifica o número de precessões de spin na segunda metade do instrumento, levando a um estado de rotação final diferente (ver Figura 1A). Isso é detectado experimentalmente como uma perda de polarização, como será mostrado mais tarde neste artigo. Para mais detalhes sobre a técnica NSE, o leitor é encaminhado para documentos técnicos dedicados42,43,44,45.
Aqui, uma descrição simplificada é apresentada para fornecer uma estimativa aproximada do comprimento e escalas de tempo acessíveis com a NSE. As escalas de comprimento são determinadas pela faixa de transferências de vetores de onda alcançáveis, Q = 4π sin φ/λ, onde 2φ é o ângulo de dispersão e λ é o comprimento de onda de nêutrons. Pode-se ver que Q é definido pela faixa de comprimento de onda e a extensão da rotação do segundo braço do espectrômetro (ver Figura 1A). Uma faixa típica de Qnos espectrômetros NSE é ~0,02-2 Å-146,47, e até 0,01-4 Å-1 com atualizações recentes48,49, correspondentes a escalas espaciais de ~1-600 Å. Por outro lado, a escala de tempo acessível é calculada a partir do ângulo total de precessão (ou fase) adquirido pelo nêutron dentro das bobinas de precessão magnética, e é encontrado como50: 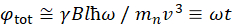 . Nesta expressão, t é o tempo de Fourier definido como
. Nesta expressão, t é o tempo de Fourier definido como 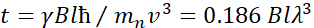 , onde está a razão
, onde está a razão 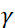 gyrommagnética de nêutrons,
gyrommagnética de nêutrons, 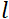 é o comprimento da bobina, e
é o comprimento da bobina, e 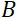 é a força do campo magnético da bobina. Vale ressaltar que o tempo de Fourier é uma quantidade estritamente dependente da geometria do instrumento, força do campo magnético e comprimento de onda de nêutrons. Por exemplo, usando nêutrons de comprimento de onda
é a força do campo magnético da bobina. Vale ressaltar que o tempo de Fourier é uma quantidade estritamente dependente da geometria do instrumento, força do campo magnético e comprimento de onda de nêutrons. Por exemplo, usando nêutrons de comprimento de onda 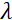 = 8 Å e configurações de instrumentos de = 1,2 m e = 0,4 T, o tempo de Fourier é calculado para ser t ~ 50 ns. Experimentalmente, o tempo de Fourier é sintonizado alterando a corrente nas bobinas de precessão (ou seja, força do campo magnético) ou usando diferentes comprimentos de onda de nêutrons, resultando em escalas típicas de tempo NSE de ~ 1 ps a 100 ns. No entanto, os upgrades recentes nos espectrômetros NSE permitiram o acesso a tempos mais longos do Fourier, até ~400 ns no espectrômetro J-NSE-Phoenix no espectrômetro Heinz Maier-Leibnitz Zentrum51 e no espectrômetro SNS-NSE no Oak Ridge National Lab48, e até ~1.000 ns no espectrômetro IN15 NSE no Institut Laue-Langevin (III)49.
= 8 Å e configurações de instrumentos de = 1,2 m e = 0,4 T, o tempo de Fourier é calculado para ser t ~ 50 ns. Experimentalmente, o tempo de Fourier é sintonizado alterando a corrente nas bobinas de precessão (ou seja, força do campo magnético) ou usando diferentes comprimentos de onda de nêutrons, resultando em escalas típicas de tempo NSE de ~ 1 ps a 100 ns. No entanto, os upgrades recentes nos espectrômetros NSE permitiram o acesso a tempos mais longos do Fourier, até ~400 ns no espectrômetro J-NSE-Phoenix no espectrômetro Heinz Maier-Leibnitz Zentrum51 e no espectrômetro SNS-NSE no Oak Ridge National Lab48, e até ~1.000 ns no espectrômetro IN15 NSE no Institut Laue-Langevin (III)49.
Além do acesso direto ao comprimento e escala de tempo da dinâmica da membrana, a NSE possui as capacidades inerentes à sensibilidade ao isótopo de nêutrons52. Especificamente, a capacidade dos nêutrons de interagir de forma diferente com os isótopos do hidrogênio, o elemento mais abundante em sistemas biológicos, resulta em uma densidade de comprimento de dispersão de nêutrons diferente,34 ou NSLD (o equivalente ao índice óptico de refração50), quando o prótium é substituído por deutério. Isso permite uma abordagem conhecida como variação de contraste, que é comumente usada para destacar características específicas da membrana ou ocultar outras — este último cenário é referido como correspondência de contraste. Uma aplicação frequente de variação/correspondência de contraste é a substituição de água (NSLD = -0,56 × 10-6 Å-2) por água pesada ou D2O (NSLD = 6,6 4 × 10-6 Å-2) para amplificar o sinal de nêutrons de membranas lipídicas protiadas (NSLD ~ 0 × 10-6 Å-2). Essa abordagem é altamente eficaz em estudos de estrutura de membrana porque a penetração de D2O na região do headgroup da membrana permite a determinação precisa das espessuras da membrana (ver Figura 2A, painel esquerdo) e da localização de diferentes subgrupos lipídes quando modelos mais sofisticados são aplicados53,54. Este artigo destaca alguns exemplos sobre o uso de variação de contraste para estudos de dinâmica coletiva em membranas biomiméticas e características de membrana selecionadas.
Aqui, a eficácia da NSE em fornecer insights únicos sobre propriedades de membrana dinâmica e funcional é ilustrada através de exemplos tangíveis de estudos NSE sobre sistemas de membrana lipídica modelo e biologicamente relevantes com ênfase na dinâmica da mesoescala em membranas autônomas, na forma de suspensões liposômicas. Para as medições NSE da dinâmica da membrana in-plane, o leitor é encaminhado para publicações dedicadas sobre espectroscopia de spin-echo de nêutrons de incidência de pastagem (GINSES)55,56 e outros estudos de pilhas de membrana multilamelaralinhadas 57,58,59,60.
Para simplificar, este artigo destaca três diferentes esquemas de deuteração de membrana ilustrados em uma formação de domínio bem estudada, ou separação de fase, sistema de bicamadas lipídicas de misturas 1,2-dimyristoyl-sn-glycero-3-fosphocholina (DMPC) e 1,2-distearoyl-sn-glycero-3-phosphocholine (DSPC) misturas61,62. Os dois lipídios são caracterizados por uma incompatibilidade em seu comprimento da cadeia de hidrocarbonetos (14 carbonos/cauda em DMPC vs 18 carbonos/cauda em DSPC) e sua temperatura de transição gel-fluido (Tm, DMPC = 23 °C vs Tm, DSPC = 55 °C). Isso resulta na separação lateral de fases nas membranas DMPC:DSPC a temperaturas entre as temperaturas de transição superior e inferior da mistura63. Os esquemas de deuteração aqui considerados são escolhidos para demonstrar os diferentes modos dinâmicos acessíveis nas medidas NSE em membranas lipoesômicas, ou seja, flutuações de dobra, flutuações de espessura e flutuações seletivas de dobra/espessura dos domínios laterais. Todas as composições lipídicas são relatadas para bicamadas DMPC:DSPC preparadas a uma fração toupeira de 70:30, utilizando variantes protiadas e semeadas comercialmente disponíveis de DMPC e DSPC. Todas as etapas de preparação da amostra são baseadas em 4 mL de suspensão liposômica, em D2O, com concentração lipídica de 50 mg/mL, para uma massa lipídica total de Mtot = 200 mgs por amostra.
