La comprensione delle membrane cellulari e della loro funzione si è notevolmente evoluta negli ultimi decenni. La precedente visione delle membrane cellulari come doppi strati lipidici passivi che definiscono i confini cellulari e le proteine di membrana1 si è gradualmente trasformata in un modello dinamico in cui i doppi strati lipidici svolgono un ruolo importante nella regolazione dei processi biologici vitali, tra cui la segnalazione cellulare, lo scambio molecolare e la funzione proteica – per citarne alcuni2,3,4,5,6. Questa consapevolezza che le membrane cellulari sono altamente dinamiche, costantemente in fase di rimodellamento e ridistribuzione molecolare, ha sollecitato esplorazioni scientifiche oltre le strutture di equilibrio delle membrane7,8,9. Di conseguenza, sono stati sviluppati diversi approcci per studiare le varie modalità dinamiche nelle membrane lipidiche biologiche e bioispira. Ad oggi, la maggior parte di questi studi si è concentrata principalmente sui moti molecolari diffusivi10,11, 12,13 e sulle fluttuazioni macroscopiche della forma14,15,16,lasciando un vuoto significativo nella comprensione della dinamica intermedia della membrana, cioè fluttuazioni collettive di assemblaggi lipidici costituiti da pochi 10-100 di molecole lipidiche. Queste dinamiche si verificano su scale di lunghezza da poche decine a poche 100 Å e su scale temporali di sub-ns a poche centinaia di ns (vedi Figura 1), qui indicate come scale mesoscopiche. È infatti su queste scale che l’attività biologica chiave si svolge a livello di membrana17. Ciò include il germogliamento virale18,il gating del canale19e le interazioni membrana-proteina20. È anche importante sottolineare che il panorama energetico delle proteine di membrana21,22 mostra che i cambiamenti conformazionali nelle proteine – necessari per il loro ruolo regolatore – avvengono sulle scale temporali ns23 delle fluttuazioni collettive della membrana, sottolineando ulteriormente l’importanza della dinamica mesoscopica nella funzione biologica delle membrane cellulari e dei loro analoghi bioispirati20. Questo documento si concentra sulle due principali modalità dinamiche mesoscopiche nelle membrane lipidiche, vale a dire, fluttuazioni di flessione e fluttuazioni di spessore.
La sfida principale nel sondare direttamente queste modalità di fluttuazione è la difficoltà di accedere simultaneamente alle loro scale spaziali e temporali utilizzando metodi di spettroscopia standard. L’altra sfida è che le tecniche di contatto diretto potrebbero avere un impatto sulle stesse fluttuazioni che dovrebbero misurare16. Ciò è ulteriormente esacerbato dalla complessità composizionale e strutturale delle membrane biologiche24,25, che si traduce in caratteristiche di membrana non omogenee, tra cui la formazione del dominio lipidico26,27,28,29,30 e l’asimmetria di membrana31,32,33– che richiedono sonde selettive per comprendere la dinamica delle diverse caratteristiche della membrana. Fortunatamente, queste sfide possono essere superate con metodi di spettroscopia neutronica non invasivi, come l’eco di spin neutronico (NSE), che accedono intrinsecamente alle scale di lunghezza e tempo richieste e consentono ulteriormente studi sulle caratteristiche selettive della membrana senza modificare il loro ambiente fisico-chimico34. Infatti, negli ultimi anni la spettroscopia NSE si è evoluta in una sonda unica e potente di dinamica collettiva della membrana35. I risultati degli studi NSE sulle membrane lipidiche hanno prodotto nuove intuizioni sulle proprietà meccaniche36,37 e viscoelastiche38,39 delle membrane lipidiche e hanno gettato nuova luce sul loro potenziale ruolo nella funzione biologica40,41.
La tecnica di spettroscopia NSE si basa su un progetto di strumento interferometrico, proposto per la prima volta da Mezei42, utilizzando una serie di pinne di spin e bobine magnetiche per controllare la precessione dello spin neutronico mentre i neutroni attraversano lo strumento. Il design si basa sulla specchiatura magnetica degli elementi del campo magnetico rispetto alla posizione del campione (Figura 1A). Ciò implica che in assenza di scambio di energia tra il neutrone e il campione, il neutrone esegue lo stesso numero di precessione di spin, in direzioni opposte, nella prima e nella seconda metà dello strumento (si noti la π-flipper tra le due bobine di precessione). Di conseguenza, lo stato di spin finale del neutrone rimane invariato rispetto allo stato iniziale – un fenomeno denominato spin-eco (vedi neutrone trasparente in Figura 1A). Tuttavia, quando il neutrone interagisce energeticamente con il campione, lo scambio di energia modifica il numero di precessione di spin nella seconda metà dello strumento, portando ad un diverso stato di spin finale (vedi Figura 1A). Questo è sperimentalmente rilevato come una perdita di polarizzazione, come verrà mostrato più avanti in questo articolo. Per maggiori dettagli sulla tecnica NSE, si rimanda al lettore gli articoli tecnici dedicati42,43,44,45.
Qui viene presentata una descrizione semplificata per fornire una stima approssimativa della lunghezza e delle scale temporali accessibili con NSE. Le scale di lunghezza sono determinate dall’intervallo di trasferimenti di motovettori realizzabili, Q = 4π sin θ/λ, dove 2θ è l’angolo di diffusione e λ è la lunghezza d’onda del neutrone. Si può vedere che Q è impostato dall’intervallo di lunghezze d’onda e dall’estensione della rotazione del secondo braccio dello spettrometro (vedi Figura 1A). Un tipico intervallo Qsugli spettrometri NSE è ~ 0,02-2 Å-146,47e fino a 0,01-4 Å-1 con recenti aggiornamenti48,49, corrispondenti a scale spaziali di ~ 1-600 Å. D’altra parte, la scala temporale accessibile è calcolata dall’angolo di precessione totale (o fase) acquisito dal neutrone all’interno delle bobine di precessione magnetica, e si trova ad essere50: 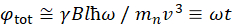 . In questa espressione, t è il tempo di Fourier definito come
. In questa espressione, t è il tempo di Fourier definito come 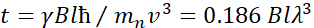 , dove è il rapporto
, dove è il rapporto 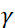 giromagnetico neutronico,
giromagnetico neutronico, 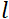 è la lunghezza della bobina e è la forza del campo
è la lunghezza della bobina e è la forza del campo 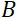 magnetico della bobina. Vale la pena sottolineare che il tempo di Fourier è una quantità che dipende strettamente dalla geometria dello strumento, dall’intensità del campo magnetico e dalla lunghezza d’onda dei neutroni. Ad esempio, usando neutroni di lunghezza d’onda
magnetico della bobina. Vale la pena sottolineare che il tempo di Fourier è una quantità che dipende strettamente dalla geometria dello strumento, dall’intensità del campo magnetico e dalla lunghezza d’onda dei neutroni. Ad esempio, usando neutroni di lunghezza d’onda 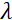 = 8 Å e impostazioni dello strumento di = 1,2 m e = 0,4 T, il tempo di Fourier è calcolato per essere t ~ 50 ns. Sperimentalmente, il tempo di Fourier viene regolato modificando la corrente nelle bobine di precessione (cioè l’intensità del campo magnetico) o utilizzando diverse lunghezze d’onda dei neutroni, con conseguente scala temporale tipica di NSE da ~ 1 ps a 100 ns. Tuttavia, i recenti aggiornamenti degli spettrometri NSE hanno consentito l’accesso a tempi di Fourier più lunghi, fino a ~ 400 ns sullo spettrometro J-NSE-Phoenix presso lo spettrometro Heinz Maier-Leibnitz Zentrum51 e lo spettrometro SNS-NSE presso l’Oak Ridge National Lab48e fino a ~ 1.000 ns presso lo spettrometro NSE IN15 presso l’Institut Laue-Langevin (ILL)49.
= 8 Å e impostazioni dello strumento di = 1,2 m e = 0,4 T, il tempo di Fourier è calcolato per essere t ~ 50 ns. Sperimentalmente, il tempo di Fourier viene regolato modificando la corrente nelle bobine di precessione (cioè l’intensità del campo magnetico) o utilizzando diverse lunghezze d’onda dei neutroni, con conseguente scala temporale tipica di NSE da ~ 1 ps a 100 ns. Tuttavia, i recenti aggiornamenti degli spettrometri NSE hanno consentito l’accesso a tempi di Fourier più lunghi, fino a ~ 400 ns sullo spettrometro J-NSE-Phoenix presso lo spettrometro Heinz Maier-Leibnitz Zentrum51 e lo spettrometro SNS-NSE presso l’Oak Ridge National Lab48e fino a ~ 1.000 ns presso lo spettrometro NSE IN15 presso l’Institut Laue-Langevin (ILL)49.
Oltre all’accesso diretto alla scala di lunghezza e tempo della dinamica di membrana, NSE ha le capacità intrinseche della sensibilità isotopica dei neutroni52. In particolare, la capacità dei neutroni di interagire in modo diverso con gli isotopi dell’idrogeno, l’elemento più abbondante nei sistemi biologici, si traduce in una diversa densità di lunghezza di scattering neutronico,34 o NSLD (l’equivalente dell’indice ottico di rifrazione50), quando il prozio è sostituito dal deuterio. Ciò consente un approccio noto come variazione di contrasto, che viene comunemente utilizzato per evidenziare specifiche caratteristiche della membrana o nascondene altre – quest’ultimo scenario è indicato come corrispondenza di contrasto. Un’applicazione frequente di variazione/corrispondenza del contrasto è la sostituzione dell’acqua (NSLD = -0,56 ×10 -6 Å -2) con acqua pesante o D2O (NSLD = 6,4 × 10-6 Å-2) per amplificare il segnale neutronico dalle membrane lipidiche protiate (NSLD ~ 0 × 10-6 Å-2). Questo approccio è altamente efficace negli studi sulla struttura della membrana perché la penetrazione di D2O nella regione del gruppo di testa della membrana consente una determinazione accurata degli spessori della membrana (vedi Figura 2A,pannello di sinistra) e della posizione di diversi sottogruppi lipidici quando vengono applicati modelli più sofisticati53,54. Questo documento evidenzia alcuni esempi sull’uso della variazione di contrasto per studi di dinamica collettiva in membrane biomimetiche e caratteristiche selezionate della membrana.
Qui, l’efficacia di NSE nel fornire approfondimenti unici sulle proprietà dinamiche e funzionali della membrana è illustrata attraverso esempi tangibili di studi NSE su sistemi di membrana lipidica modello e biologicamente rilevanti con particolare attenzione alla dinamica della mesoscala nelle membrane autoportanti, sotto forma di sospensioni liposomiali. Per le misure NSE della dinamica della membrana in-plane, il lettore è rimandato a pubblicazioni dedicate sulla spettroscopia spin-eco neutronica ad incidenza radente (GINSES)55,56 e altri studi su pile di membrane multilamellari allineate57,58,59,60.
Per semplicità, questo articolo evidenzia tre diversi schemi di deuterazione della membrana illustrati su un sistema a doppio strato lipidico ben studiato di formazione di dominio, o separazione di fase, di miscele di 1,2-dimiristoil-sn-glicero-3-fosfocolina (DMPC) e 1,2-distearoil-sn-glicero-3-fosfocolina (DSPC)61,62. I due lipidi sono caratterizzati da una discrepanza nella lunghezza della catena idrocarburica (14 atomi di carbonio/coda in DMPC vs 18 atomi di carbonio/coda in DSPC) e dalla loro temperatura di transizione gel-fluido (Tm, DMPC = 23 °C vs Tm, DSPC = 55 °C). Ciò si traduce in una separazione di fase laterale nelle membrane DMPC:DSPC a temperature comprese tra le temperature di transizione superiore e inferiore della miscela63. Gli schemi di deuterazione qui considerati sono scelti per dimostrare le diverse modalità dinamiche accessibili nelle misurazioni NSE su membrane liposomiali, vale a dire, fluttuazioni di flessione, fluttuazioni di spessore e fluttuazioni selettive di flessione / spessore di domini laterali. Tutte le composizioni lipidiche sono riportate per i doppi strati DMPC:DSPC preparati a una frazione molare di 70:30, utilizzando varianti proziate e perdeuterate disponibili in commercio di DMPC e DSPC. Tutte le fasi di preparazione del campione si basano su 4 mL di sospensione liposomiale, in D2O, con una concentrazione lipidica di 50 mg/mL, per una massa lipidica totale di Mtot = 200 mg per campione.
