L’assorbimento opportunistico di nutrienti extracellulari intratumorali (cioè) è stato recentemente nominato un segno distintivo chiave per il metabolismo del cancro1. Uno di questi importanti nutrienti è l’ATP, in quanto la concentrazione di ieATP è10 3 e10 4 volte superiore a quella che si trova nei tessuti normali, nell’intervallo da diverse centinaia di μM a bassi mM2,3,4,5. Come molecola chiave di energia e segnalazione, l’ATP svolge un ruolo centrale nel metabolismo cellulare nelle cellule cancerose e sane6,7,8. L’ATP extracellulare non è solo coinvolto nella crescita delle cellule tumorali, ma promuove anche la resistenza ai farmaci9. Funzioni precedentemente non riconosciute dell’ATP, come l’attività idrotropica, sono state recentemente identificate, implicando così il coinvolgimento dell’ATP in malattie come l’Alzheimer10. In effetti, sembra che la nostra comprensione dell’ATP e delle sue funzioni nelle cellule tumorali, nelle cellule sane e in altre cellule malate sia tutt’altro che completa. Tuttavia, a causa dell’instabilità dell’ATP e degli alti tassi di turnover nelle cellule, è tecnicamente difficile monitorare il movimento dell’ATP attraverso la membrana cellulare e nella cellula.
Per affrontare questo problema e soddisfare le esigenze di questa area di ricerca, è stato sviluppato un metodo in cui l’ATP fluorescente non idrolizzabile (NHF-ATP) (Figura 1) è stato utilizzato come surrogato per visualizzare l’internalizzazione dell’ATP e osservare la localizzazione spaziale intracellulare dell’ATP internalizzato, sia in vitro che in vivo11,12 . NHF-ATP ha dimostrato di sostituire l’ATP endogeno per studiare il movimento dell’ATP attraverso le membrane cellulari animali, sia nelle linee cellulari tumorali che nel tessuto tumorale umano xenotrapiantato su topi immunodeficienti11,12. Inoltre, la somministrazione di inibitori della macropinocitosi alle cellule ha bloccato l’internalizzazione dell’eATP, suggerendo che l’assorbimento intracellulare di eATP comporta un meccanismo macropinocitotico9,11,12. Questo protocollo consente il colabeling immunobased contro proteine cellulari specifiche e quindi l’identificazione di quale tipo di cellula interiorizza NHF-ATP. Utilizzando xenotrapianti tumorali in vivo e microscopia ad alta risoluzione, NHF-ATP può essere visualizzato spazialmente attraverso il campione di tessuto e anche all’interno di una singola cellula. Questi metodi consentono anche analisi quantitative, come la percentuale di assorbimento cellulare, il numero di vescicole macropinocitotiche e la cinetica di internalizzazione. Questo articolo descrive in dettaglio come NHF-ATP, lavorando da solo o insieme adextrans13, 14,15,16fluorescenti endocitosi-traccianti, può essere utilizzato in diversi contesti sperimentali per studiare l’internalizzazione e la localizzazione intracellulare dell’ATP, a seguito dell’internalizzazione nelle cellule.
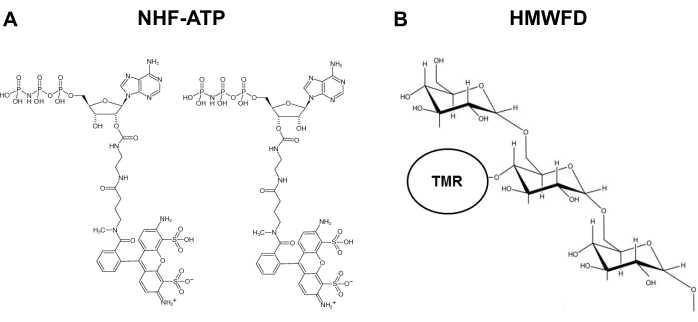
Figura 1: Strutture di ATP fluorescente non idrolizzabile e tetrametilrodramina etichettate come destrano fluorescente ad alto peso molecolare. (A) Struttura di NHF-ATP. (B) Rappresentazione schematica dell’HMWFD. Abbreviazioni: ATP = adenosina trifosfato; NHF-ATP = ATP fluorescente non idrolizzabile; TMR = tetrametilrodramina; HMWFD = destrano fluorescente ad alto peso molecolare. Fare clic qui per visualizzare una versione più grande di questa figura.
