Det opportunistiske opptaket av intratumorale ekstracellulære (dvs.) næringsstoffer har nylig blitt kalt et sentralt kjennetegn for kreftmetabolisme1. Et av disse viktige næringsstoffene er ATP, da konsentrasjonen av ieATP er 103 og 104 ganger høyere enn den som finnes i normalt vev, i området flere hundre μM til lav mM2,3,4,5. Som et viktig energi- og signalmolekyl spiller ATP en sentral rolle i cellulær metabolisme i kreft og sunne celler6,7,8. Ekstracellulær ATP er ikke bare involvert i kreftcellevekst, men det fremmer også legemiddelresistens9. Tidligere ukjente funksjoner av ATP, som hydrotropisk aktivitet, har nylig blitt identifisert, og dermed implisert ATP-involvering i sykdommer som Alzheimers10. Faktisk ser det ut til at vår forståelse av ATP og dens funksjoner i kreftceller, sunne celler og andre syke celler er langt fra komplett. På grunn av ATPs ustabilitet og høye omsetningshastigheter i celler er det imidlertid teknisk utfordrende å overvåke ATPs bevegelse over cellemembranen og inn i cellen.
For å løse dette problemet og fylle behovet for dette forskningsområdet, ble det utviklet en metode der ikke-hydroolysert fluorescerende ATP (NHF-ATP) (figur 1) ble brukt som surrogat for å visualisere internaliseringen av ATP og observere den intracellulære romlige lokaliseringen av internalisert ATP, både in vitro og in vivo11,12 . NHF-ATP har vist seg å erstatte endogen ATP for å undersøke ATP-bevegelse på tvers av dyrecellemembraner, både i kreftcellelinjer og i humant tumorvev xenograftert på immunodeficient mus11,12. Videre, administrere makropinocytosehemmere til celler blokkert eATP internalisering, noe som tyder på at intracellulært opptak av eATP innebærer en makropinocytotisk mekanisme9,11,12. Denne protokollen tillater immunobased colabeling mot cellespesifikke proteiner og dermed identifisering av hvilken celletype som internaliserer NHF-ATP. Ved hjelp av in vivo tumor xenografts og høyoppløselig mikroskopi kan NHF-ATP visualiseres romlig over vevsprøven og til og med i en enkelt celle. Disse metodene tillater også kvantitativ analyse, for eksempel prosentandelen av cellulært opptak, antall makropinocytotiske vesikler og internaliseringskinetikk. Dette dokumentet beskriver i detalj hvordan NHF-ATP, som arbeider alene eller sammen med endokytose-tracer fluorescerende dextrans13,14,15,16, kan brukes i forskjellige eksperimentelle omgivelser for å studere ATPs internalisering og intracellulær lokalisering, etter internalisering i celler.
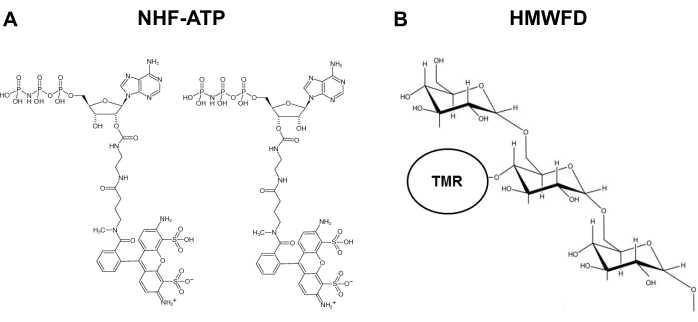
Figur 1: Strukturer av ikke-hydroolyzable fluorescerende ATP og tetrametylrhodamin merket høymolekylær fluorescerende dextran. (A) Struktur av NHF-ATP. (B) Skjematisk representasjon av HMWFD. Forkortelser: ATP = adenosin tripfosfat; NHF-ATP = ikke-hydroolyzable fluorescerende ATP; TMR = tetrametylrhodamin; HMWFD = fluorescerende dekstran med høy molekylvekt. Klikk her for å se en større versjon av denne figuren.
