中子是一种无电荷的大质量粒子,多年来已成功用于探测从基础物理学到生物学1等各个领域的样品。对于生物应用,小角中子散射,非弹性中子散射以及中子晶体学和反射计被广泛使用2,3,4。非弹性中子散射提供了动力学的集合平均测量,而不需要特定的标记本身,并且信号质量不依赖于大小或蛋白质5。对于模拟细胞内培养基的所研究蛋白质,例如氘代细菌裂解物甚至体内3,6,7,可以使用高度复杂的环境进行测量。可以使用不同的实验装置来研究动力学,即i)飞行时间访问sub-ps-ps动力学,ii)反向散射-访问ps-ns动力学,以及iii)自旋回波访问从ns到数百ns的动力学。中子反向散射利用布拉格定律 2d sinθ = nλ,其中 d 是晶体中平面之间的距离,θ 是散射角,n 是散射阶数,λ 是波长。使用晶体向检测器进行反向散射可以实现高分辨率的能量,通常为~0.8μeV。为了测量能量交换,可以使用携带反向散射晶体的多普勒驱动器来定义和调整入射中子波长8,9,10(图1),或者可以使用飞行时间设置,但代价是能量分辨率降低11。
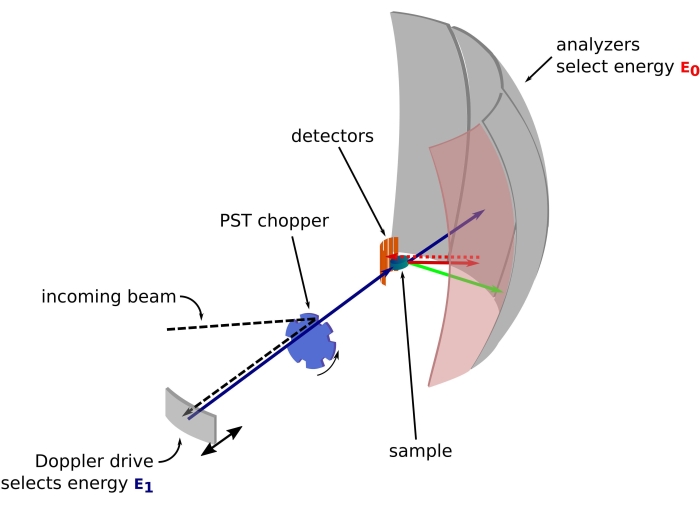
图 1:带多普勒驱动器的中子背散射光谱仪的草图。入射光束撞击相空间变换(PST)斩波器42,这增加了样品位置的通量。然后通过多普勒驱动器将其反向散射到样品,多普勒驱动器选择能量E1(青色箭头)。然后中子被样品散射(箭头的颜色表示不同的能量),由Si 111晶体制成的分析仪只会反向散射具有特定能量E0的中子(此处为红色箭头)。因此,动量传递q是从探测器阵列上中子的检测位置获得的,能量转移是从差值E1– E0获得的。PST产生的中子脉冲的预期飞行时间用于丢弃直接散射到探测器管的中子的信号。缩写:PST = 相空间变换。请点击此处查看此图的大图。
对于背散射光谱,富含氢质子的样品(如蛋白质)信号的主要贡献来自非相干散射,其散射强度 Sinc(q, ω) 由方程 (1)12 表示
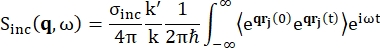 (1)
(1)
其中σinc是所考虑元素的非相干截面,k’是散射波矢量的范数,k是入射波矢量的范数,q(= k – k’)是动量传递,r j(t)是原子j在时间t的位置矢量,ω是对应于入射中子和系统之间能量转移的频率。尖括号表示融合平均值。因此,非相干散射探测原子位置随时间的集合平均单粒子自相关性,并给出系统中所有原子和不同时间起源的自动力学平均值(集合平均值)。散射函数是中间散射函数 I(q, t) 在时间上的傅里叶变换,可以看作是方程 (2) 所示的范霍夫相关函数在空间中的傅里叶变换:
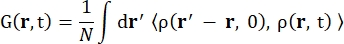 (二)
(二)
其中 ρ(r,t) 是在位置 r 和时间t 13 处找到原子的概率密度。
对于菲克扩散过程,自扩散函数在散射函数中进行双傅里叶变换后产生(参见方程(3)),散射函数由γ = Dq2给出的线宽洛伦兹组成。
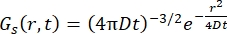 (三)
(三)
开发了更复杂的模型并发现有用,例如Singwi和Sjölander用于ps-ns内部蛋白质动力学14的跳跃扩散模型或Sears用于水合水15,16,17的旋转模型。
在法国格勒诺布尔ILL的中子背散射(NBS)仪器IN16B8,9上(补充图S1),蛋白质常用的设置由Si 111晶体组成,用于分析仪的Si 111晶体,带有用于调整入射波长的多普勒驱动器(补充图S2A),从而可以访问动量转移范围~0.2 Å-1 < q < ~2 Å-1和-30μeV的能量转移范围< 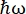 < 30 μeV – 对应于从几ps到几ns的时间尺度和几Å的距离。此外,IN16B还提供了执行弹性和非弹性固定窗口扫描(E/IFWS)10的可能性,其中包括固定能量传输的数据采集。由于在处理中子时通量受到限制,E/IFWS允许在一次能量转移中最大化通量,从而减少获得令人满意的信噪比所需的采集时间。最近的选择是反向散射和飞行时间光谱仪(BATS)模式11,它允许测量广泛的能量传输(例如,-150μeV< <150μeV),具有比多普勒驱动器更高的通量,但代价是能量分辨率较低(补充图S2B)。
< 30 μeV – 对应于从几ps到几ns的时间尺度和几Å的距离。此外,IN16B还提供了执行弹性和非弹性固定窗口扫描(E/IFWS)10的可能性,其中包括固定能量传输的数据采集。由于在处理中子时通量受到限制,E/IFWS允许在一次能量转移中最大化通量,从而减少获得令人满意的信噪比所需的采集时间。最近的选择是反向散射和飞行时间光谱仪(BATS)模式11,它允许测量广泛的能量传输(例如,-150μeV< <150μeV),具有比多普勒驱动器更高的通量,但代价是能量分辨率较低(补充图S2B)。
中子散射的一个重要特性是,inc σ非相干截面的氢值比氘高40倍,对于生物样品中常见的其他元素可以忽略不计。因此,可以使用氘代缓冲液研究液体环境中蛋白质的动力学,并且粉末状态允许研究用D 2 O水合的氢化蛋白粉的蛋白质内部动力学,或研究用H2O水合的全氘化蛋白粉的水合水。在液态下,中子背散射通常允许同时访问蛋白质的质心自扩散(Fickian型扩散)及其内部动力学。后者是骨干和侧链运动,通常由所谓的跳跃扩散模型或其他模型描述 3,18.在氢化蛋白粉中,不存在蛋白质扩散,只需要对内部动力学进行建模。对于水合水,水分子的平移和旋转运动的贡献对动量传递q具有不同的依赖性,这使得它们在数据分析过程中得以区分17。
本文通过研究蛋白质来说明中子背散射方法,这些蛋白质被发现能够展开,聚集成由β链堆栈组成的规范形式 – 所谓的交叉β模式19,20 – 并形成细长的纤维。这就是所谓的淀粉样蛋白聚集,由于其在阿尔茨海默氏症或帕金森病等神经退行性疾病中的核心作用而被广泛研究21,22。淀粉样蛋白的研究还受到它们可以发挥的功能作用23,24或其在开发新型生物材料方面的高潜力的动机25。淀粉样蛋白聚集的物理化学决定因素尚不清楚,尽管在过去几年中取得了巨大进展,但没有淀粉样蛋白聚集的一般理论可用21,26。
淀粉样蛋白聚集意味着蛋白质结构和稳定性随时间的变化,其研究自然意味着动力学,与蛋白质构象稳定性、蛋白质功能和蛋白质能量景观27 有关。动力学通过对最快运动28的熵贡献与特定状态的稳定性直接相关,蛋白质功能可以通过从光敏蛋白29 的sub-ps到域运动的ms的各种时间尺度上的运动来维持,这可以通过皮秒-纳秒动力学30来促进。
将介绍两个使用中子背散射光谱研究淀粉样蛋白的例子,一个在液态研究蛋白质动力学,另一个在水合粉末状态下研究水合水动力学。第一个示例涉及实时将溶菌酶聚集成μm大小的球体(称为颗粒)5,第二个示例比较人类蛋白质tau31的天然和聚集状态下的水动力学。
溶菌酶是一种参与免疫防御的酶,由129个氨基酸残基组成。溶菌酶可以在pD为10.5和温度为90°C的氘代缓冲液中形成颗粒。 通过中子散射,我们发现溶菌酶质心扩散系数的时间演变遵循硫黄素T荧光(用于监测淀粉样蛋白交叉β模式形成的荧光探针32)的单指数动力学,表明形成颗粒上层结构和交叉β模式以相同的速率在单个步骤中发生。此外,内部动力学在整个聚集过程中保持不变,这可以通过在NBS仪器上无法观察到的快速构象变化来解释,或者可以通过聚集时蛋白质内能没有显着变化来解释。
人类蛋白tau是一种固有的无序蛋白(IDP),由441个氨基酸组成,用于所谓的2N4R亚型,其主要参与阿尔茨海默病33。在全氘化蛋白tau粉末上使用中子反向散射,我们发现在纤维状态下水合水动力学增加,更多的水分子群经历平移运动。结果表明,水合水熵的增加可能驱动tau的淀粉样蛋白颤动。
