הנייטרון הוא חלקיק מסיבי וחסר מטען ששימש בהצלחה לאורך השנים לבדיקת דגימות בתחומים שונים, מפיזיקה בסיסית ועד ביולוגיה1. עבור יישומים ביולוגיים, פיזור נייטרונים בזווית קטנה, פיזור נייטרונים בלתי אלסטי, קריסטלוגרפיה ורפלקטומטריה של נייטרונים נמצאים בשימוש נרחב 2,3,4. פיזור נייטרונים קשיח מספק מדידה ממוצעת של הדינמיקה ללא צורך בתיוג ספציפי כשלעצמו, ואיכות אות שאינה תלויה בגודל או בחלבון5. המדידה יכולה להיעשות באמצעות סביבה מורכבת מאוד עבור החלבון הנחקר המחקה את התווך התוך-תאי, כגון ליזט חיידקי או אפילו in vivo 3,6,7. ניתן להשתמש במערכי ניסוי שונים כדי לחקור את הדינמיקה, כלומר i) גישה של זמן טיסה לדינמיקות sub-ps-ps, ii) גישה המעניקה פיזור לאחור לדינמיקות ps-ns, ו-iii) גישה נותנת ספין-הד לדינמיקה מ-ns למאות ns. פיזור אחורי של נייטרונים עושה שימוש בחוק בראג 2d sinθ = nλ, כאשר d הוא המרחק בין מישורים בגביש, θ זווית הפיזור, n סדר הפיזור ו-λ אורך הגל. השימוש בגבישים לפיזור לאחור לכיוון הגלאים מאפשר השגת רזולוציה גבוהה באנרגיה, בדרך כלל ~0.8 μeV. כדי למדוד את חילופי האנרגיה, משתמשים בכונן דופלר הנושא גביש בפיזור לאחור כדי להגדיר ולכוונן את אורך גל הנייטרונים הנכנס 8,9,10 (איור 1), או שניתן להשתמש במערך זמן טיסה במחיר של ירידה ברזולוציית האנרגיה 11.
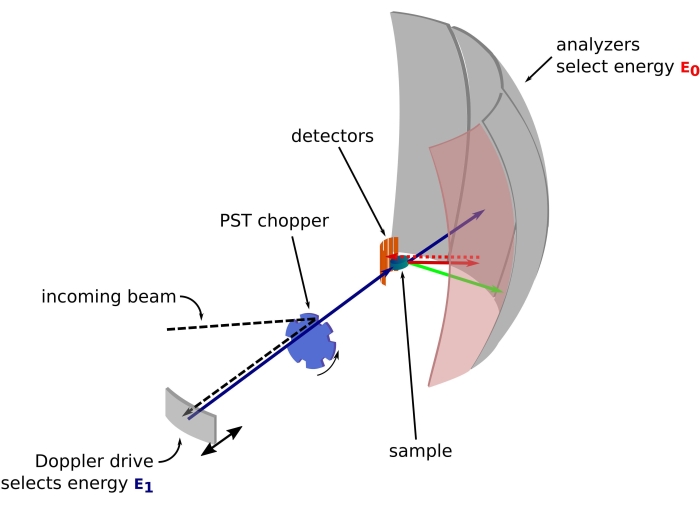
איור 1: שרטוט של ספקטרומטר פיזור לאחור של נייטרונים עם כונן דופלר. הקרן הנכנסת פוגעת במסוק טרנספורמציית חלל הפאזה (PST)42, המגדיל את השטף במיקום הדגימה. לאחר מכן הוא מפוזר לאחור לכיוון הדגימה על ידי כונן דופלר, אשר בוחר אנרגיה E1 (חץ ציאן). לאחר מכן הנייטרונים מפוזרים על ידי הדגימה (עם אנרגיות שונות המיוצגות על ידי צבע החיצים) והאנלייזרים, העשויים מגבישי Si 111, יפזרו לאחור נייטרונים רק עם אנרגיה ספציפית E0 (חיצים בצבע אדום כאן). לפיכך, העברת התנע q מתקבלת מהמיקום שזוהה של הנייטרון על מערך הגלאים, והעברת האנרגיה מתקבלת מההפרש E1– E0. זמן הטיסה הצפוי לפעימת הנייטרונים המופקת על ידי ה-PST משמש להשלכת האות מהנייטרונים המפוזרים ישירות לעבר צינורות הגלאי. קיצור: PST = טרנספורמציה של מרחב פאזה. אנא לחץ כאן כדי להציג גרסה גדולה יותר של איור זה.
עבור ספקטרוסקופיית פיזור לאחור, התרומה העיקרית לאות מדגימות עשירות בפרוטוני מימן, כגון חלבונים, מגיעה מפיזור לא קוהרנטי, שעבורו עוצמת הפיזור Sinc(q, ω) מוצגת על ידי Eq (1)12
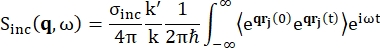 (1)
(1)
כאשר σinc הוא חתך הרוחב הלא קוהרנטי של היסוד הנחשב, k’ הוא הנורמה של וקטור הגל המפוזר, k הנורמה של וקטור הגל הנכנס, q (= k – k’) העברת התנע, r j(t) וקטור המיקום של אטום j בזמן t, ו-ω התדר המתאים למעבר האנרגיה בין הנייטרון הנכנס למערכת. הסוגריים הזוויתיים מציינים את ממוצע ההרכב. לפיכך, פיזור לא קוהרנטי בוחן את המתאם העצמי הממוצע של חלקיק בודד של מיקומי אטומים עם זמן ונותן את הדינמיקה העצמית הממוצעת על פני כל האטומים במערכת ומקורות זמן שונים (ממוצע אנסמבל). פונקציית הפיזור היא התמרת פורייה בזמן של פונקציית פיזור הביניים I(q, t), שניתן לראותה כהתמרת פורייה במרחב של פונקציית מתאם ואן הוב המוצגת על ידי Eq (2):
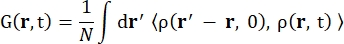 (2)
(2)
כאשר ρ(r,t) היא צפיפות ההסתברות למציאת אטום במיקום r ובזמן t 13.
עבור תהליך דיפוזיה פיקיאנית, פונקציית הדיפוזיה העצמית נוצרת (ראו Eq (3)) לאחר התמרת פורייה כפולה בפונקציית פיזור המורכבת מלורנציאן של רוחב קו הנתון על ידי γ = Dq2.
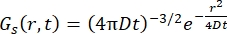 (3)
(3)
מודלים מתוחכמים יותר פותחו ונמצאו שימושיים כגון מודל דיפוזיית הקפיצה על ידי Singwi ו- Sjölander עבור ps-ns דינמיקה פנימית של חלבונים14 או מודל הסיבוב על ידי סירס עבור מי הידרציה15,16,17.
במכשיר פיזור אחורי של נייטרונים (NBS) IN16B8,9 ב-ILL, גרנובל, צרפת (איור משלים S1), מערך נפוץ עם חלבונים מורכב מגבישי Si 111 עבור האנלייזרים עם כונן דופלר לכוונון אורך הגל הנכנס (איור משלים S2A), ובכך נותן גישה לטווח העברת התנע ~0.2 Å-1 < q < ~2 Å-1 וטווח העברת אנרגיה של –30 μeV < 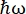 < 30 μeV – מתאים לטווחי זמן הנעים בין כמה ps לכמה ns ומרחקים של כמה Å. בנוסף, IN16B מציע את האפשרות לבצע סריקות גמישות ובלתי אלסטיות של חלונות קבועים (E/IFWS)10, הכוללות איסוף נתונים בהעברת אנרגיה קבועה. מכיוון שהשטף מוגבל בעבודה עם נייטרונים, E/IFWS מאפשר למקסם את השטף להעברת אנרגיה אחת, ובכך מקצר את זמן הרכישה הדרוש לקבלת יחס אות לרעש מספק. אפשרות עדכנית יותר היא מצב ספקטרומטר פיזור לאחור וזמן טיסה (BATS)11, המאפשר מדידה של מגוון רחב של העברות אנרגיה, (למשל, -150 μeV < < 150 μeV), עם שטף גבוה יותר מאשר עם כונן דופלר, אך במחיר של רזולוציית אנרגיה נמוכה יותר (איור משלים S2B).
< 30 μeV – מתאים לטווחי זמן הנעים בין כמה ps לכמה ns ומרחקים של כמה Å. בנוסף, IN16B מציע את האפשרות לבצע סריקות גמישות ובלתי אלסטיות של חלונות קבועים (E/IFWS)10, הכוללות איסוף נתונים בהעברת אנרגיה קבועה. מכיוון שהשטף מוגבל בעבודה עם נייטרונים, E/IFWS מאפשר למקסם את השטף להעברת אנרגיה אחת, ובכך מקצר את זמן הרכישה הדרוש לקבלת יחס אות לרעש מספק. אפשרות עדכנית יותר היא מצב ספקטרומטר פיזור לאחור וזמן טיסה (BATS)11, המאפשר מדידה של מגוון רחב של העברות אנרגיה, (למשל, -150 μeV < < 150 μeV), עם שטף גבוה יותר מאשר עם כונן דופלר, אך במחיר של רזולוציית אנרגיה נמוכה יותר (איור משלים S2B).
תכונה חשובה של פיזור נייטרונים היא שחתך הרוחב הלא קוהרנטי σinc הוא בעל ערך גבוה פי 40 עבור מימן מאשר עבור דאוטריום, והוא זניח עבור יסודות אחרים הנמצאים בדרך כלל בדגימות ביולוגיות. לכן, ניתן לחקור את הדינמיקה של חלבונים בסביבה נוזלית באמצעות מאגר deuterated, ומצב האבקה מאפשר לחקור דינמיקה פנימית של חלבונים עם אבקת חלבון מוקשה hydrated עם D 2 O, או המחקר של מים הידרציה עבוראבקת חלבון perdeuterated hydrated עם H2O. במצב נוזלי, פיזור אחורי של נייטרונים מאפשר בדרך כלל גישה בו זמנית לדיפוזיה העצמית של מרכז המסה של חלבונים (דיפוזיה מסוג פיקיאן) ולדינמיקה הפנימית שלהם. אלה האחרונים הם תנועות עמוד השדרה ושרשרת הצד המתוארות בדרך כלל על ידי מה שנקרא מודל דיפוזיה קפיצה או אחרים 3,18. באבקות חלבון מוקשה, דיפוזיית החלבון נעדרת ויש למדל רק דינמיקה פנימית. עבור מי הידרציה, התרומות של תנועות תרגומיות וסיבוביות של מולקולות מים מציגות תלות שונה בהעברת התנע q, המאפשרת את ההבחנה ביניהן בתהליך ניתוח הנתונים17.
מאמר זה מדגים את שיטת הפיזור לאחור של נייטרונים באמצעות מחקר של חלבונים שנמצאו מסוגלים להתפתח, להצטבר לצורה קנונית המורכבת מערימות של β-גדילים – מה שמכונה תבנית β צולבת19,20 – וליצור סיבים מוארכים. זהו מה שנקרא צבירה עמילואיד, אשר נחקר בהרחבה בשל תפקידו המרכזי בהפרעות נוירודגנרטיביות כגון אלצהיימר או פרקינסון21,22. המחקר של חלבוני העמילואיד מונע גם על ידי התפקיד הפונקציונלי שהם יכולים למלא 23,24 או הפוטנציאל הגבוה שלהם לפיתוח ביו-חומרים חדשים25. הדטרמיננטים הפיזיקוכימיים של צבירת העמילואיד עדיין אינם ברורים, ואין תיאוריה כללית של צבירת עמילואיד, למרות התקדמות עצומה במהלך השנים האחרונות21,26.
צבירת עמילואיד מרמזת על שינויים במבנה החלבון וביציבותו עם הזמן, שהמחקר שלהם מרמז באופן טבעי על דינמיקה, הקשורה ליציבות קונפורמציה של חלבונים, תפקוד חלבונים ונוף אנרגיית חלבון27. דינמיקה קשורה ישירות ליציבות של מצב מסוים באמצעות התרומה האנטרופית עבור התנועות המהירות ביותר28, ותפקוד החלבונים יכול להתקיים על ידי תנועות בסקאלות זמן שונות, החל מ-sub-ps עבור חלבונים רגישים לאור29 ועד ms עבור תנועות תחום, אשר ניתן להקל על ידי דינמיקה פיקו-שנייה-ננו-שנייה30.
יוצגו שתי דוגמאות לשימוש בספקטרוסקופיית פיזור לאחור של נייטרונים לחקר חלבוני עמילואיד, אחת במצב נוזלי לחקר דינמיקה של חלבונים ואחת במצב אבקה רוויית לחות לחקר דינמיקת מי הידרציה. הדוגמה הראשונה נוגעת לצבירה של ליזוזים לכדורים בגודל מיקרומטר (הנקראים חלקיקים) ואחריה בזמן אמת5, והשנייה השוואה של דינמיקת מים במצבים טבעיים ומצטברים של החלבון האנושי טאו31.
ליזוזים הוא אנזים המעורב בהגנה החיסונית ומורכב משאריות של 129 חומצות אמינו. ליזוזים יכול ליצור חלקיקים בחיץ מפורק ב pD של 10.5 ובטמפרטורה של 90 ° C. עם פיזור נייטרונים, הראינו כי התפתחות הזמן של מקדם הדיפוזיה של מרכז המסה של ליזוזים עוקבת אחר הקינטיקה המעריכית היחידה של תיאופלבין T פלואורסצנטי (בדיקה פלואורסצנטית המשמשת לניטור היווצרות תבניות β צולבות עמילואיד32), מה שמצביע על כך שהיווצרות מבני על חלקיקיים ותבניות β צולבות מתרחשות בשלב אחד עם אותו קצב. יתר על כן, הדינמיקה הפנימית נשארה קבועה לאורך כל תהליך הצבירה, אשר ניתן להסביר או על ידי שינוי קונפורמציה מהיר שלא ניתן לצפות בו על מכשירי NBS, או על ידי היעדר שינוי משמעותי באנרגיה הפנימית של החלבונים בעת הצבירה.
החלבון האנושי טאו הוא חלבון בעל הפרעה מהותית (IDP) המורכב מ-441 חומצות אמינו עבור מה שמכונה איזופורם 2N4R, אשר מעורב באופן בולט במחלת אלצהיימר33. באמצעות פיזור לאחור של נייטרונים על אבקות של חלבון טאו מפורר, הראינו כי דינמיקת מי הידרציה מוגברת במצב הסיבים, כאשר אוכלוסייה גבוהה יותר של מולקולות מים עוברות תנועות תרגומיות. התוצאה מצביעה על כך שעלייה באנטרופיית מי הידרציה עשויה להניע את פרפור העמילואיד של טאו.
