Neutronen er en ladningsfri og massiv partikel, der med succes er blevet brugt gennem årene til at undersøge prøver på forskellige områder fra grundlæggende fysik til biologi1. Til biologiske anvendelser anvendes småvinkel neutronspredning, uelastisk neutronspredning og neutronkrystallografi og reflektometri i vid udstrækning 2,3,4. Uelastisk neutronspredning giver en ensemble-gennemsnitlig måling af dynamikken uden at kræve specifik mærkning i sig selv og en signalkvalitet, der ikke afhænger af størrelsen eller proteinet5. Målingen kan udføres ved hjælp af et meget komplekst miljø for det undersøgte protein, der efterligner det intracellulære medium, såsom et deutereret bakterielt lysat eller endda in vivo 3,6,7. Forskellige eksperimentelle opsætninger kan bruges til at studere dynamikken, nemlig i) time-of-flight-giver adgang til sub-ps-ps-dynamik, ii) backscattering-giver adgang til ps-ns-dynamik og iii) spin-ekko-giver adgang til dynamik fra ns til hundredvis af ns. Neutron backscattering gør brug af Braggs lov 2d sinθ = nλ, hvor d er afstanden mellem planer i en krystal, θ spredningsvinklen, n spredningsrækkefølgen og λ bølgelængden. Brugen af krystaller til backscattering mod detektorerne giver mulighed for at opnå en høj opløsning i energi, typisk ~ 0,8 μeV. For at måle energiudvekslingen bruges enten et Doppler-drev, der bærer en krystal i backscattering, til at definere og indstille den indkommende neutronbølgelængde 8,9,10 (figur 1), eller en time-of-flight-opsætning kan bruges på bekostning af et fald i energiopløsning 11.
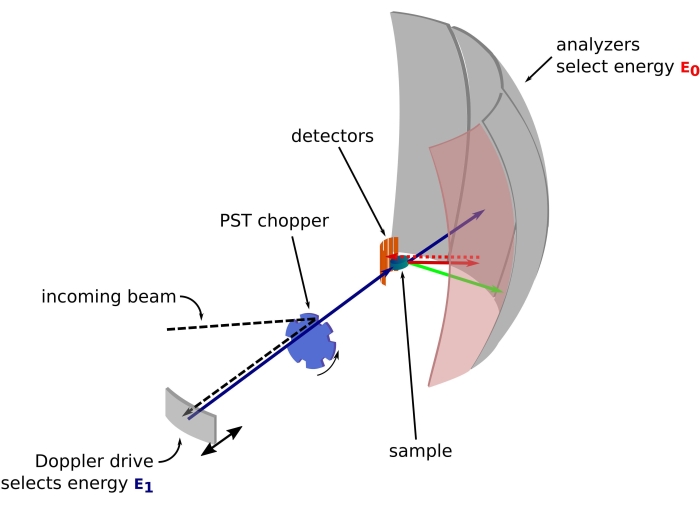
Figur 1: Skitse af et neutron backscattering spektrometer med et Doppler-drev. Den indkommende stråle rammer faserumstransformationen (PST) chopper42, hvilket øger fluxen ved prøvepositionen. Det spredes derefter tilbage mod prøven af Doppler-drevet, som vælger en energi E1 (cyanpil). Neutronerne spredes derefter af prøven (med forskellige energier repræsenteret af pilenes farve), og analysatorerne, der er lavet af Si 111-krystaller, vil kun backscatter neutroner med en bestemt energi E0 (rødfarvede pile her). Derfor opnås momentumoverførslen q fra neutronens detekterede position på detektorarrayet, og energioverførslen opnås fra forskellen E1– E0. Den forventede flyvetid for neutronpulsen produceret af PST bruges til at kassere signalet fra neutronerne spredt direkte mod detektorrørene. Forkortelse: PST = fase rumtransformation. Klik her for at se en større version af denne figur.
For backscattering-spektroskopi kommer hovedbidraget til signalet fra hydrogenprotonrige prøver, såsom proteiner, fra usammenhængende spredning, for hvilken spredningsintensiteten Sinc(q, ω) er vist ved Eq (1)12
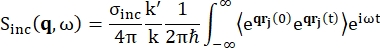 (1)
(1)
Hvor σinc er det usammenhængende tværsnit af det betragtede element, er k’ normen for den spredte bølgevektor, k normen for den indkommende bølgevektor, q (= k – k’) momentumoverførslen, r j (t) positionsvektoren for atom j på tidspunktet t og ω frekvensen svarende til energioverførslen mellem den indkommende neutron og systemet. Vinkelbeslagene angiver ensemblegennemsnittet. Derfor undersøger usammenhængende spredning den ensemble-gennemsnitlige enkeltpartikel-selvkorrelation af atompositioner med tiden og giver selvdynamikken i gennemsnit over alle atomer i systemet og forskellige tidsoprindelser (ensemblegennemsnit). Spredningsfunktionen er Fouriertransformationen i tid af den mellemliggende spredningsfunktion I(q, t), der kan ses som Fouriertransformationen i rummet af van Hove-korrelationsfunktionen vist ved Eq (2):
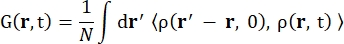 (2)
(2)
Hvor ρ(r,t) er sandsynlighedstætheden for at finde et atom i position r og tid t 13.
For en fickiansk diffusionsproces resulterer selvdiffusionsfunktionen (se Eq (3)) efter en dobbelt Fourier-transformation i en spredningsfunktion bestående af en Lorentzian med linjebredde givet ved γ = Dq2.
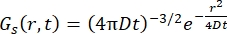 (3)
(3)
Mere sofistikerede modeller blev udviklet og fundet nyttige, såsom springdiffusionsmodellen af Singwi og Sjölander til ps-ns interne proteindynamik14 eller rotationsmodellen af Sears til hydreringsvand15,16,17.
På neutron backscattering (NBS) instrumentet IN16B 8,9 ved ILL, Grenoble, Frankrig (supplerende figur S1), består en opsætning, der almindeligvis anvendes med proteiner, af Si 111-krystaller til analysatorerne med et Doppler-drev til tuning af den indkommende bølgelængde (supplerende figur S2A), hvilket giver adgang til momentumoverførselsområdet ~ 0,2 Å-1 < q < ~ 2 Å-1 og energioverførselsområdet på –30 μeV < 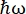 < 30 μeV – svarende til tidsskalaer fra nogle få ps til nogle få ns og afstande på nogle få Å. Derudover giver IN16B mulighed for at udføre elastiske og uelastiske fastvinduesscanninger (E/IFWS)10, som omfatter dataindsamling ved en fast energioverførsel. Da fluxen er begrænset, når man arbejder med neutroner, tillader E/IFWS maksimering af fluxen for en energioverførsel, hvilket reducerer den anskaffelsestid, der er nødvendig for at opnå et tilfredsstillende signal-støj-forhold. En nyere mulighed er backscattering og time-of-flight spektrometer (BATS) mode11, som muliggør måling af en lang række energioverførsler (f.eks. -150 μeV < < 150 μeV) med en højere flux end med Doppler-drevet, men på bekostning af en lavere energiopløsning (supplerende figur S2B).
< 30 μeV – svarende til tidsskalaer fra nogle få ps til nogle få ns og afstande på nogle få Å. Derudover giver IN16B mulighed for at udføre elastiske og uelastiske fastvinduesscanninger (E/IFWS)10, som omfatter dataindsamling ved en fast energioverførsel. Da fluxen er begrænset, når man arbejder med neutroner, tillader E/IFWS maksimering af fluxen for en energioverførsel, hvilket reducerer den anskaffelsestid, der er nødvendig for at opnå et tilfredsstillende signal-støj-forhold. En nyere mulighed er backscattering og time-of-flight spektrometer (BATS) mode11, som muliggør måling af en lang række energioverførsler (f.eks. -150 μeV < < 150 μeV) med en højere flux end med Doppler-drevet, men på bekostning af en lavere energiopløsning (supplerende figur S2B).
En vigtig egenskab ved neutronspredning er, at det usammenhængende tværsnit σinc har en 40 gange højere værdi for hydrogen end for deuterium og er ubetydelig for andre grundstoffer, der almindeligvis findes i biologiske prøver. Derfor kan dynamikken af proteiner i et flydende miljø studeres ved hjælp af en deutereret buffer, og pulvertilstanden muliggør undersøgelse af enten proteinets indre dynamik med hydrogeneret proteinpulver hydreret med D 2 Oeller undersøgelsen af hydreringsvand til perdeutereret proteinpulver hydreret med H2O. I flydende tilstand tillader neutron backscattering typisk samtidig adgang til massecentrets selvdiffusion af proteiner (Fickian-type diffusion) og deres interne dynamik. Sidstnævnte er rygrads- og sidekædebevægelser, der normalt beskrives af den såkaldte springdiffusionsmodel eller andre 3,18. I hydrogeneret proteinpulver er proteindiffusionen fraværende, og kun intern dynamik skal modelleres. For hydreringsvand præsenterer bidragene fra translationelle og rotationsbevægelser af vandmolekyler en anden afhængighed af momentumoverførslen q, hvilket muliggør deres sondring i dataanalyseprocessen17.
Dette papir illustrerer neutron backscattering metoden med undersøgelsen af proteiner, der viste sig at være i stand til at udfolde sig, aggregere i en kanonisk form bestående af stakke af β-strenge – det såkaldte cross-β mønster 19,20 – og danne aflange fibre. Dette er den såkaldte amyloidaggregering, som er grundigt undersøgt på grund af dens centrale rolle i neurodegenerative lidelser som Alzheimers eller Parkinsons sygdomme21,22. Undersøgelsen af amyloidproteinerne er også motiveret af den funktionelle rolle, de kan spille 23,24 eller deres store potentiale for udvikling af nye biomaterialer25. De fysisk-kemiske determinanter for amyloidaggregeringen forbliver uklare, og der findes ingen generel teori om amyloidaggregering på trods af enorme fremskridt i de seneste år21,26.
Amyloidaggregering indebærer ændringer i proteinstruktur og stabilitet med tiden, hvis undersøgelse naturligt indebærer dynamik, knyttet til proteinkonformationsstabilitet, proteinfunktion og proteinenergilandskab27. Dynamik er direkte knyttet til stabiliteten af en bestemt tilstand gennem det entropiske bidrag til de hurtigste bevægelser28, og proteinfunktionen kan opretholdes af bevægelser på forskellige tidsskalaer fra sub-ps for lysfølsomme proteiner29 til ms for domænebevægelser, hvilket kan lettes af picosekund-nanosekund dynamik30.
To eksempler på anvendelse af neutron backscattering spektroskopi til at studere amyloidproteiner vil blive præsenteret, et i flydende tilstand for at studere proteindynamik og et i hydratiseret pulvertilstand for at studere hydreringsvanddynamik. Det første eksempel vedrører aggregering af lysozym i μm-store kugler (kaldet partikler) efterfulgt i realtid5, og det andet en sammenligning af vanddynamik i indfødte og aggregerede tilstande af det humane protein tau31.
Lysozym er et enzym involveret i immunforsvaret og består af 129 aminosyrerester. Lysozym kan danne partikler i deutereret buffer ved pD på 10,5 og ved en temperatur på 90 °C. Med neutronspredning viste vi, at tidsudviklingen af lysozyms center-of-mass diffusionskoefficient følger den enkelte eksponentielle kinetik af thioflavin T-fluorescens (en fluorescerende sonde, der bruges til at overvåge dannelsen af amyloid cross-β mønstre32), hvilket indikerer, at formationspartikeloverbygningerne og tvær-β mønstre forekommer i et enkelt trin med samme hastighed. Desuden forblev den interne dynamik konstant gennem hele aggregeringsprocessen, hvilket kan forklares enten ved en hurtig konformationsændring, der ikke kan observeres på NBS-instrumenter, eller ved fraværet af signifikant ændring i proteinintern energi ved aggregering.
Det humane protein tau er et iboende forstyrret protein (IDP) bestående af 441 aminosyrer til den såkaldte 2N4R-isoform, som især er involveret i Alzheimers sygdom33. Ved hjælp af neutron backscattering på pulvere af perdeutereret protein tau viste vi, at hydreringsvanddynamikken øges i fibertilstanden, med en højere population af vandmolekyler, der gennemgår translationelle bevægelser. Resultatet tyder på, at en stigning i hydreringsvandentropi kan drive amyloidflimmer af tau.
