Neutronen är en laddningsfri och massiv partikel som framgångsrikt har använts genom åren för att undersöka prover inom olika områden från grundläggande fysik till biologi1. För biologiska tillämpningar används neutronspridning med liten vinkel, inelastisk neutronspridning och neutronkristallografi och reflektometri i stor utsträckning 2,3,4. Inelastisk neutronspridning ger en ensemble-medelvärdesmätning av dynamiken utan att kräva specifik märkning i sig, och en signalkvalitet som inte beror på storleken eller proteinet5. Mätningen kan göras med hjälp av en mycket komplex miljö för proteinet som studeras som efterliknar det intracellulära mediet, såsom ett deutererat bakteriellt lysat eller till och med in vivo 3,6,7. Olika experimentella uppställningar kan användas för att studera dynamiken, nämligen i) time-of-flight-giving access to sub-ps-ps dynamics, ii) backscattering-giving access to ps-ns dynamics, och iii) spin-echo-giving access to dynamics from ns to hundreds ns. Neutronåterspridning använder Braggs lag 2d sinθ = nλ, där d är avståndet mellan plan i en kristall, θ spridningsvinkeln, n spridningsordningen och λ våglängden. Användningen av kristaller för bakåtspridning mot detektorerna möjliggör en hög upplösning i energi, typiskt ~ 0,8 μeV. För att mäta energiutbytet används antingen en dopplerdrivenhet som bär en kristall i backscattering för att definiera och ställa in den inkommande neutronvåglängden 8,9,10 (figur 1), eller en flygtidsinställning kan användas på bekostnad av en minskning av energiupplösningen 11.
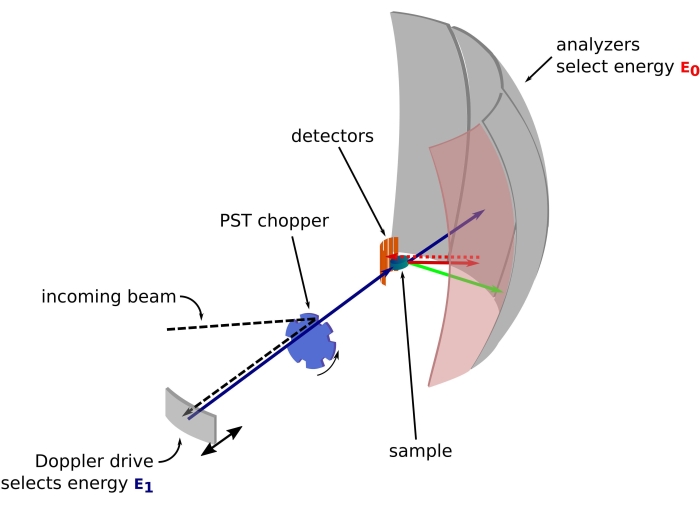
Figur 1: Skiss av en neutronspridningsspektrometer med dopplerdrift. Den inkommande strålen träffar fasrumstransformationen (PST) chopper42, vilket ökar flödet vid provpositionen. Den sprids sedan bakåt mot provet av dopplerdrivningen, som väljer en energi E1 (cyanpil). Neutronerna sprids sedan av provet (med olika energier representerade av pilarnas färg) och analysatorerna, gjorda av Si 111-kristaller, kommer bara att sprida neutroner med en specifik energi E0 (rödfärgade pilar här). Därför erhålls momentöverföringen q från neutronens detekterade position på detektormatrisen och energiöverföringen erhålls från skillnaden E1– E0. Den flygtid som förväntas för neutronpulsen som produceras av PST används för att kassera signalen från neutronerna spridda direkt mot detektorrören. Förkortning: PST = fasrymdtransformation. Klicka här för att se en större version av denna figur.
För backscatteringspektroskopi kommer det huvudsakliga bidraget till signalen från väteprotonrika prover, såsom proteiner, från inkoherent spridning, för vilken spridningsintensiteten Sinc (q, ω) visas av Eq (1) 12
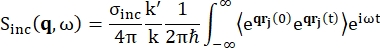 (1)
(1)
Därσ inc är det osammanhängande tvärsnittet av det element som beaktas, är k’ normen för den spridda vågvektorn, k normen för den inkommande vågvektorn, q (= k – k’) momentöverföringen, r j (t) positionsvektorn för atom j vid tiden t och ω frekvensen som motsvarar energiöverföringen mellan den inkommande neutronen och systemet. Vinkelparenteserna anger ensemblemedelvärdet. Därför undersöker inkoherent spridning ensemblemedelvärdet av enpartikels självkorrelation av atompositioner med tiden och ger självdynamiken i genomsnitt över alla atomer i systemet och olika tidsursprung (ensemblemedelvärde). Spridningsfunktionen är Fouriertransformen i tiden av den mellanliggande spridningsfunktionen I(q, t), som kan ses som Fouriertransformen i rummet av van Hove-korrelationsfunktionen som visas av Eq (2):
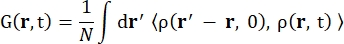 (2)
(2)
Där ρ(r,t) är sannolikhetstätheten för att hitta en atom vid position r och tiden t 13.
För en fickiansk diffusionsprocess resulterar självdiffusionsfunktionen (se Eq (3)) efter en dubbel Fouriertransform i en spridningsfunktion bestående av en Lorentzian med linjebredd given av γ = Dq2.
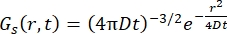 (3)
(3)
Mer sofistikerade modeller utvecklades och befanns användbara såsom hoppdiffusionsmodellen av Singwi och Sjölander för ps-ns interna proteindynamik14 eller rotationsmodellen av Sears för hydratiseringsvatten15,16,17.
På neutronspridningsinstrumentet IN16B 8,9 vid ILL, Grenoble, Frankrike (kompletterande figur S1) består en inställning som vanligtvis används med proteiner av Si 111-kristaller för analysatorerna med en dopplerdrivning för att ställa in den inkommande våglängden (kompletterande figur S2A), vilket ger tillgång till momentöverföringsområdet ~ 0,2 Å-1 < q < ~ 2 Å-1 och energiöverföringsområdet –30 μeV < 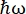 < 30 μeV motsvarande tidsskalor från några ps till några ns och avstånd på några Å. Dessutom erbjuder IN16B möjligheten att utföra elastiska och oelastiska skanningar med fasta fönster (E/IFWS)10, vilket inkluderar datainsamling vid en fast energiöverföring. Eftersom flödet är begränsat vid arbete med neutroner möjliggör E/IFWS maximering av flödet för en energiöverföring, vilket minskar förvärvstiden som behövs för att uppnå ett tillfredsställande signal-brusförhållande. Ett nyare alternativ är backscattering och time-of-flight spectrometer (BATS) mode11, som möjliggör mätning av ett brett spektrum av energiöverföringar, (t.ex. -150 μeV < < 150 μeV), med ett högre flöde än med dopplerdriften, men på bekostnad av en lägre energiupplösning (kompletterande figur S2B).
< 30 μeV motsvarande tidsskalor från några ps till några ns och avstånd på några Å. Dessutom erbjuder IN16B möjligheten att utföra elastiska och oelastiska skanningar med fasta fönster (E/IFWS)10, vilket inkluderar datainsamling vid en fast energiöverföring. Eftersom flödet är begränsat vid arbete med neutroner möjliggör E/IFWS maximering av flödet för en energiöverföring, vilket minskar förvärvstiden som behövs för att uppnå ett tillfredsställande signal-brusförhållande. Ett nyare alternativ är backscattering och time-of-flight spectrometer (BATS) mode11, som möjliggör mätning av ett brett spektrum av energiöverföringar, (t.ex. -150 μeV < < 150 μeV), med ett högre flöde än med dopplerdriften, men på bekostnad av en lägre energiupplösning (kompletterande figur S2B).
En viktig egenskap hos neutronspridning är att det inkoherenta tvärsnittet σinc har ett 40 gånger högre värde för väte än för deuterium och är försumbar för andra element som vanligtvis finns i biologiska prover. Därför kan dynamiken hos proteiner i en flytande miljö studeras med hjälp av en deutererad buffert, och pulvertillståndet möjliggör studier av antingen proteinets inre dynamik med hydrerat proteinpulver hydratiserat med D2O, eller studien av hydratiseringsvatten för perdeutererat proteinpulver hydratiserat medH2O. I flytande tillstånd tillåter neutronåterspridning typiskt samtidig åtkomst till masscentrets självdiffusion av proteiner (diffusion av fickian-typ) och deras interna dynamik. De senare är ryggrads- och sidokedjerörelser som vanligtvis beskrivs av den så kallade hoppdiffusionsmodellen eller andra 3,18. I hydrerade proteinpulver saknas proteindiffusionen och endast intern dynamik behöver modelleras. För hydratiseringsvatten uppvisar bidragen från translationella och roterande rörelser av vattenmolekyler ett annat beroende av momentöverföringen q, vilket möjliggör deras distinktion i dataanalysprocessen17.
Denna artikel illustrerar neutronspridningsmetoden med studier av proteiner som visade sig kunna utvecklas, aggregera till en kanonisk form bestående av staplar av β-strängar – det så kallade kors-β mönstret19,20 – och bilda långsträckta fibrer. Detta är den så kallade amyloidaggregeringen, som studeras omfattande på grund av dess centrala roll i neurodegenerativa sjukdomar som Alzheimers eller Parkinsons sjukdomar21,22. Studien av amyloidproteinerna motiveras också av den funktionella roll de kan spela 23,24 eller deras höga potential för utveckling av nya biomaterial25. De fysikalisk-kemiska determinanterna för amyloidaggregeringen är fortfarande oklara, och ingen allmän teori om amyloidaggregering finns tillgänglig, trots enorma framsteg under de senaste åren21,26.
Amyloidaggregering innebär förändringar i proteinstruktur och stabilitet med tiden, vars studie naturligtvis innebär dynamik, kopplad till proteinkonformationsstabilitet, proteinfunktion och proteinenergilandskap27. Dynamik är direkt kopplad till stabiliteten hos ett specifikt tillstånd genom det entropiska bidraget för de snabbaste rörelserna28, och proteinfunktionen kan upprätthållas av rörelser på olika tidsskalor från sub-ps för ljuskänsliga proteiner29 till ms för domänrörelser, vilket kan underlättas av picosecond-nanosecond dynamics30.
Två exempel på att använda neutronbackspridningsspektroskopi för att studera amyloidproteiner kommer att presenteras, ett i flytande tillstånd för att studera proteindynamik och ett i hydratiserat pulvertillstånd för att studera hydratiseringsvattendynamik. Det första exemplet gäller aggregering av lysozym i μm-sfärer (kallade partiklar) följt i realtid5, och det andra en jämförelse av vattendynamik i naturliga och aggregerade tillstånd av det humana proteinet tau31.
Lysozym är ett enzym som är involverat i immunförsvaret och består av 129 aminosyrarester. Lysozym kan bilda partiklar i deutererad buffert vid pD 10,5 och vid en temperatur på 90 °C. Med neutronspridning visade vi att tidsutvecklingen av massdiffusionskoefficienten för lysozym följer den enda exponentiella kinetiken för tioflavin T-fluorescens (en fluorescerande sond som används för att övervaka bildandet av amyloidkors-β mönster32), vilket indikerar att bildningspartikelformiga överbyggnader och kors-β mönster förekommer i ett enda steg med samma hastighet. Dessutom förblev den interna dynamiken konstant under hela aggregeringsprocessen, vilket kan förklaras antingen av en snabb konformationsförändring som inte kan observeras på NBS-instrument eller av frånvaron av signifikant förändring i proteinets interna energi vid aggregering.
Det humana proteinet tau är ett egenstört protein (IDP) som består av 441 aminosyror för den så kallade 2N4R-isoformen, som är särskilt involverad i Alzheimers sjukdom33. Med hjälp av neutronåterspridning på pulver av perdeutererat protein tau visade vi att hydratiseringsvattendynamiken ökar i fibertillståndet, med en högre population av vattenmolekyler som genomgår translationella rörelser. Resultatet tyder på att en ökning av hydreringsvattenentropi kan driva amyloidflimmer av tau.
