الطماطم (S. lycopersicum L.) هي واحدة من أهم محاصيل الخضروات في جميع أنحاء العالم ، حيث يبلغ إنتاجها 186.8 مليون طن من الفواكه السمين من 5.1 مليون هكتار في عام 20201. ينتمي إلى عائلة Solanaceae الكبيرة مع حوالي 2716 نوعا2 ، بما في ذلك العديد من المحاصيل المهمة تجاريا مثل الباذنجان والفلفل والبطاطس والتبغ. الطماطم المزروعة هي نوع ثنائي الصيغة الصبغية (2n = 2x = 24) بحجم جينوم يبلغ حوالي 900 ميجا بايت3. لفترة طويلة ، تم بذل جهد كبير نحو تدجين الطماطم وتكاثرها عن طريق اختيار السمات المرغوبة من Solanum spp. هناك أكثر من 5000 مدخل طماطم مدرج في مركز موارد علم الوراثة للطماطم ويتم تخزين أكثر من 80000 مادة وراثية من الطماطم في جميع أنحاء العالم4. نبات الطماطم معمر في الدفيئة وينتشر بالبذور. تتكون بذرة الطماطم الناضجة من ثلاث حجرات رئيسية: جنين كامل النمو ، السويداء الخلوي المتبقي ، ومعطف البذور الصلب 5,6 (الشكل 1 أ). بعد الإخصاب المزدوج ، يسبق تطور السويداء من النوع الخلوي تطور الزيجوت. في ~ 5-6 أيام بعد الإزهار (DAF) ، لوحظ الجنين ثنائي الخلية لأول مرة عندما يتكون السويداء من ستة إلى ثمانية نوى7. في Solanum pimpinellifolium ، يقترب الجنين من حجمه النهائي بعد 20 DAF ، والبذور قابلة للإنبات بعد 32 DAF8. مع تطور الجنين ، يتم امتصاص السويداء تدريجيا وتبقى كمية صغيرة فقط من السويداء في البذور. يتكون السويداء المتبقي من السويداء micropylar المحيطة بطرف الجذر ، والسويداء الجانبي في بقية البذور 9,10. تم تطوير طبقة البذور الخارجية من البشرة الخارجية السميكة والخشنة للتكامل ، ومع الطبقات الميتة من بقايا التكامل ، فإنها تشكل قشرة صلبة لحماية الجنين والسويداء5.
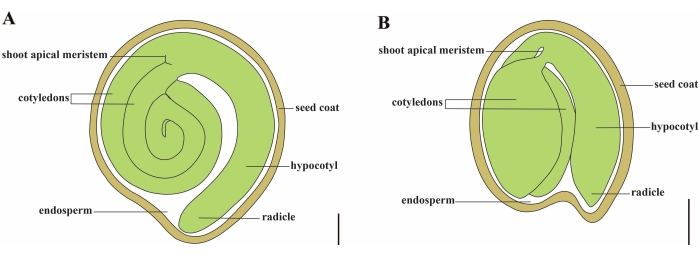
الشكل 1: تمثيل تخطيطي لبذرة ناضجة في Solanum lycopersicum و Arabidopsis thaliana. أ: التشريح الطولي لبذور الطماطم الناضجة. (ب) التشريح الطولي لبذرة أرابيدوبسيس ناضجة. يبلغ حجم بذور الطماطم الناضجة حوالي 70 مرة أكبر من بذور أرابيدوبسيس. قضبان المقياس = (أ) 400 ميكرومتر ، (ب) 100 ميكرومتر. الرجاء النقر هنا لعرض نسخة أكبر من هذا الشكل.
يعتمد إنتاج بذور الطماطم عالية الجودة على التنسيق بين الجنين والسويداء ومكونات بذور الأم11. يتطلب تشريح الجينات والشبكات الرئيسية في تطوير البذور تسجيلا ظاهريا عميقا وكاملا للبذور الطافرة. يتم تطبيق تقنيات التضمين التقليدية ، مثل القسم شبه الرقيق وقسم البارافين ، على نطاق واسع على بذور الطماطم لمراقبة الهياكل المحلية والدقيقة للجنين12،13،14،15. ومع ذلك ، فإن تحليل تطور البذور من المقاطع الرقيقة عادة ما يكون شاقا ويفتقر إلى الدقة المكانية للمحور z. بالمقارنة ، تعد إزالة الأنسجة طريقة سريعة وفعالة لتحديد المرحلة التنموية لعيوب الجنين التي من المرجح أن تحدث16. تقلل طريقة المقاصة من عتامة الأنسجة الداخلية عن طريق تجانس معامل الانكسار مع واحد أو أكثر من العوامل الكيميائية الحيوية16. تسمح إزالة الأنسجة الكاملة بمراقبة بنية الأنسجة النباتية دون تدمير سلامتها ، وأصبح الجمع بين تقنية المقاصة والتصوير ثلاثي الأبعاد حلا مثاليا للحصول على معلومات حول التشكل والحالة التنموية لعضو النبات17,18. على مر السنين ، تم استخدام تقنيات إزالة البذور في أنواع نباتية مختلفة ، بما في ذلك Arabidopsis thaliana و Hordeum vulgare و Beta vulgaris19،20،21،22،23. من بين هذه ، كانت تقنية إزالة البويضات الكاملة نهجا فعالا لدراسة تطور بذور Arabidopsis ، نظرا لصغر حجمها ، و 4-5 طبقات من خلية معطف البذور ، والسويداء من النوع النووي24,25. مع التحديث المستمر لمخاليط المقاصة المختلفة ، مثل ظهور محلول هوير26 ، تم تصوير الهياكل الداخلية لبويضة الشعير بدرجة عالية من الوضوح على الرغم من أن السويداء يشكل الجزء الأكبر من البذور. يمكن ملاحظة التطور الجنيني لبنجر السكر عن طريق التطهير مع المعالجة الفراغية والتليين بحمض الهيدروكلوريك19. ومع ذلك ، على عكس الأنواع المذكورة أعلاه ، لم يتم الإبلاغ عن الملاحظات الجنينية عن طريق بروتوكولات الإزالة في بذور الطماطم. هذا يمنع التحقيق التفصيلي في التطور الجنيني والبذور للطماطم.
يستخدم هيدرات الكلورال بشكل شائع كمحلول تطهير يسمح بعرض الأنسجة والخلايا المغمورة على مستويات بصرية مختلفة ، ويحافظ بشكل كبير على الخلايا أو مكونات الأنسجة27،28،29. تم استخدام بروتوكول المقاصة القائم على هيدرات الكلورال بنجاح لإزالة البذور بالكامل لمراقبة الجنين والسويداء في Arabidopsis21,28. ومع ذلك ، فإن محلول المقاصة هذا ليس فعالا في إزالة بذور الطماطم ، والتي هي أكثر نفاذية من بذور Arabidopsis. تشمل الحواجز المادية: (1) يحتوي تكامل الطماطم على ما يقرب من 20 طبقة خلية من 3 إلى 15 DAF 30,31 ، (2) السويداء الطماطم من النوع الخلوي ، وليس من النوعالنووي 32 ، و (3) بذور الطماطم أكبر بحوالي 70 مرة في الحجم 33,34 و (4) تنتج كميات كبيرة من صمغ معطف البذور ، مما يمنع تغلغل الكواشف المطهرة ويؤثر على تصور خلايا الجنين.
لذلك ، يقدم هذا التقرير طريقة محسنة للمقاصة القائمة على هيدرات الكلورال لإزالة بذور الطماطم بالكامل في مراحل مختلفة ، مما يسمح بالتصوير العميق لعملية تطور الجنين (الشكل 2).
