Tomate (S. lycopersicum L.) ist eine der wichtigsten Gemüsekulturen der Welt, mit einer Produktion von 186,8 Millionen Tonnen fleischigen Früchten von 5,1 Millionen Hektar im Jahr 20201. Es gehört zur großen Familie der Solanaceae mit etwa 2.716 Arten2, darunter viele kommerziell wichtige Kulturen wie Auberginen, Paprika, Kartoffeln und Tabak. Die Kulturtomate ist eine diploide Art (2n = 2x = 24) mit einer Genomgröße von ca. 900Mb3. Seit langem werden große Anstrengungen unternommen, um Tomatendomestizierung und -züchtung zu züchten, indem wünschenswerte Merkmale aus wilden Solanum spp. Es gibt über 5.000 Tomatenakzessionen, die im Tomato Genetics Resource Center aufgeführt sind, und mehr als 80.000 Keimplasma von Tomaten werden weltweit gelagert4. Die Tomatenpflanze ist mehrjährig im Gewächshaus und vermehrt sich durch Samen. Ein reifer Tomatensamen besteht aus drei Hauptkompartimenten: einem ausgewachsenen Embryo, einem zellulären Endosperm und einer harten Samenschale 5,6 (Abbildung 1A). Nach doppelter Befruchtung geht die Entwicklung von zellulärem Endosperm der Entwicklung von Zygoten voraus. Bei ~5-6 Tagen nach der Blüte (DAF) wird erstmals ein zweizelliger Proembryo beobachtet, wenn das Endosperm aus sechs bis acht Kernen besteht7. Bei Solanum pimpinellifolium nähert sich der Embryo nach 20 DAF seiner endgültigen Größe, und die Samen sind nach 32 DAF8 für die Keimung geeignet. Wenn sich der Embryo entwickelt, wird das Endosperm allmählich absorbiert und nur eine kleine Menge Endosperm verbleibt im Samen. Das restliche Endosperm besteht aus mikropylarem Endosperm, das die Radikelspitze umgibt, und seitlichem Endosperm im Rest des Samens 9,10. Die äußere Samenhülle wird aus verdickter und verholzter äußerer Epidermis des Integuments entwickelt, und mit den abgestorbenen Schichten von Integumentresten bilden sie eine harte Schale, um den Embryo und das Endosperm zu schützen5.
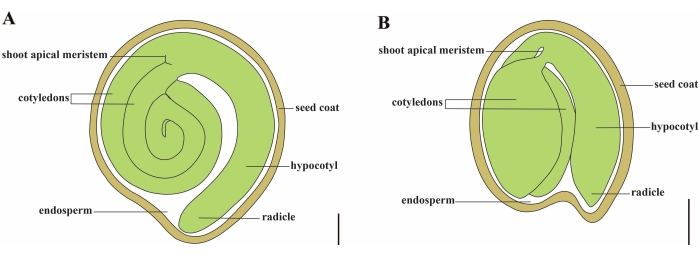
Abbildung 1: Schematische Darstellung eines reifen Samens in Solanum lycopersicum und Arabidopsis thaliana. (A) Längsanatomie eines reifen Tomatensamens. (B) Längsanatomie eines reifen Arabidopsis-Samens. Ein reifer Tomatensamen ist etwa 70-mal größer als ein Arabidopsis-Samen. Maßstabsbalken = (A) 400 μm, (B) 100 μm. Bitte klicken Sie hier, um eine größere Version dieser Abbildung zu sehen.
Die Produktion hochwertiger Tomatensamen hängt von der Koordination zwischen dem Embryo, dem Endosperm und den mütterlichen Samenbestandteilen ab11. Die Analyse von Schlüsselgenen und Netzwerken in der Samenentwicklung erfordert eine tiefe und vollständige phänotypische Aufzeichnung von mutierten Samen. Konventionelle Einbettungsschnitttechniken, wie der Halbdünnschliff und der Paraffinschnitt, werden häufig auf Tomatensamen angewendet, um die lokalen und feineren Strukturen des Embryoszu beobachten 12,13,14,15. Die Analyse der Seed-Entwicklung aus dünnen Schnitten ist jedoch in der Regel mühsam und es fehlt die räumliche Auflösung der z-Achse. Im Vergleich dazu ist die Gewebereinigung eine schnelle und effiziente Methode, um das Entwicklungsstadium von embryonalen Defekten zu bestimmen, die am wahrscheinlichsten auftreten16. Das Clearing-Verfahren verringert die Undurchsichtigkeit des inneren Gewebes durch Homogenisierung des Brechungsindex mit einem oder mehreren biochemischen Mitteln16. Die gesamte Gewebereinigung ermöglicht die Beobachtung einer pflanzlichen Gewebestruktur, ohne ihre Integrität zu zerstören, und die Kombination von Clearing-Technologie und dreidimensionaler Bildgebung ist zu einer idealen Lösung geworden, um Informationen über die Morphologie und den Entwicklungszustand eines Pflanzenorgans zu erhalten17,18. Im Laufe der Jahre wurden Samenreinigungstechniken bei verschiedenen Pflanzenarten eingesetzt, darunter Arabidopsis thaliana, Hordeum vulgare und Beta vulgaris 19,20,21,22,23. Unter diesen war die Whole-mount-Eizellenreinigungstechnologie ein effizienter Ansatz zur Untersuchung der Samenentwicklung von Arabidopsis, aufgrund ihrer geringen Größe, 4-5 Schichten der Samenhüllenzelle und des nuklearen Endosperms24,25. Mit der kontinuierlichen Aktualisierung verschiedener Clearing-Mischungen, wie dem Auftauchen der Hoyer-Lösung26, wurden die inneren Strukturen der Gerstenzelle mit einem hohen Grad an Klarheit abgebildet, obwohl ihr Endosperm den Großteil der Samen ausmacht. Die Embryogenese von Zuckerrüben kann durch Reinigung in Kombination mit Vakuumbehandlung und Erweichung mit Salzsäurebeobachtet werden 19. Im Gegensatz zu den oben genannten Arten wurden jedoch keine embryologischen Beobachtungen durch Clearing-Protokolle in Tomatensamen berichtet. Dies verhindert eine detaillierte Untersuchung der Embryonal- und Samenentwicklung von Tomaten.
Chloralhydrat wird üblicherweise als Reinigungslösung verwendet, die es ermöglicht, die eingetauchten Gewebe und Zellen auf verschiedenen optischen Ebenen darzustellen und die Zellen oder Gewebekomponenten im Wesentlichen zu erhalten 27,28,29. Das auf Chloralhydrat basierende Clearing-Protokoll wurde erfolgreich für die gesamte Reinigung von Samen verwendet, um den Embryo und das Endosperm von Arabidopsis21,28 zu beobachten. Diese Clearing-Lösung ist jedoch nicht effizient bei der Reinigung von Tomatensamen, die undurchlässiger sind als Arabidopsis-Samen. Zu den physischen Barrieren gehören: (1) das Tomatenintegument hat fast 20 Zellschichten bei 3 bis 15 DAF 30,31, (2) das Tomatenendosperm ist zellulär, nicht vom Kerntyp 32, und (3) Tomatensamen sind etwa70-mal größer in der Größe33,34 und (4) produzieren große Mengen an Samenmantelschleimstoffen, die das Eindringen von Reinigungsreagenzien blockieren und die Visualisierung von Embryozellen beeinträchtigen.
Daher stellt dieser Bericht eine optimierte Chloralhydrat-basierte Clearing-Methode für die vollständige Reinigung von Tomatensamen in verschiedenen Stadien vor, die eine tiefe Bildgebung des Embryo-Entwicklungsprozesses ermöglicht (Abbildung 2).
