Томат (S. lycopersicum L.) является одной из важнейших овощных культур во всем мире, с объемом производства 186,8 млн. тонн мясистых фруктов с 5,1 млн. га в 2020 году1. Он принадлежит к большому семейству пасленовых, насчитывающему около 2 716 видов2, включая многие коммерчески важные культуры, такие как баклажаны, перец, картофель и табак. Культивируемый помидор является диплоидным видом (2n = 2x = 24) с размером генома около 900 Mb3. В течение долгого времени были приложены большие усилия для одомашнивания и селекции томатов путем отбора желаемых признаков из дикого Solanum spp. В Центре генетических ресурсов томатов зарегистрировано более 5000 томатов, и во всем мире хранится более 80 000 зародышевой плазмы помидоров4. Растение томат является многолетним в теплице и размножается семенами. Зрелое семя томата состоит из трех основных отсеков: полноценного эмбриона, остаточного эндосперма клеточного типа и твердой семенной оболочки 5,6 (рисунок 1А). После двойного оплодотворения развитию эндосперма клеточного типа предшествует развитие зигот. Через ~5-6 дней после цветения (DAF) впервые наблюдается двухклеточное проэмбрио, когда эндосперм состоит из шести-восьми ядер7. В Solanum pimpinellifolium эмбрион приближается к своему окончательному размеру после 20 DAF, а семена жизнеспособны для прорастания после 32 DAF8. По мере развития эмбриона эндосперм постепенно всасывается и в семени остается лишь небольшое количество эндосперма. Остаточный эндосперм состоит из микропирлярного эндосперма, окружающего кончик радика, и бокового эндосперма в остальной части семени 9,10. Наружная семенная оболочка развивается из утолщенного и одревесного наружного эпидермиса покрова, а с отмершими слоями остатков покрова они образуют твердую оболочку для защиты эмбриона и эндосперма5.
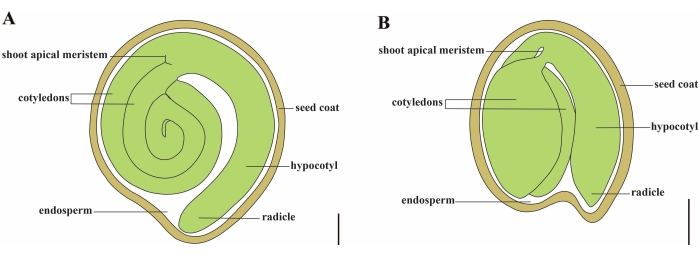
Рисунок 1: Схематическое изображение зрелого семени у Solanum lycopersicum и Arabidopsis thaliana. (A) Продольная анатомия зрелого семени томата. (B) Продольная анатомия зрелого семени Arabidopsis . Зрелое семя помидора примерно в 70 раз больше по размеру, чем семя арабидопсиса . Шкала = (A) 400 мкм, (B) 100 мкм. Пожалуйста, нажмите здесь, чтобы просмотреть увеличенную версию этого рисунка.
Производство высококачественных семян томатов зависит от координации между компонентами эмбриона, эндосперма и материнского семенного компонента11. Препарирование ключевых генов и сетей в развитии семян требует глубокой и полной фенотипической регистрации мутантных семян. Традиционные методы встраивания-сечения, такие как полутонкое сечение и парафиновое сечение, широко применяются к семенам томатов для наблюдения за местными и более тонкими структурами эмбриона 12,13,14,15. Однако анализ развития семян из тонких участков обычно трудоемкий и не имеет пространственного разрешения по оси Z. Для сравнения, очистка тканей является быстрым и эффективным методом для точного определения стадии развития дефектов эмбриона, которые, скорее всего, произойдут16. Способ очистки уменьшает непрозрачность внутренней ткани путем гомогенизации показателя преломления одним или несколькими биохимическими агентами16. Очистка всей ткани позволяет наблюдать структуру растительной ткани, не разрушая ее целостность, а сочетание технологии очистки и трехмерной визуализации стало идеальным решением для получения информации о морфологии и состоянии развития растительного органа17,18. На протяжении многих лет методы очистки семян использовались у различных видов растений, включая Arabidopsis thaliana, Hordeum vulgare и Beta vulgaris 19,20,21,22,23. Среди них технология очистки яйцеклеток с цельной горой была эффективным подходом к изучению развития семян Arabidopsis, благодаря его небольшому размеру, 4-5 слоям клетки семенной оболочки и эндосперму ядерного типа24,25. При постоянном обновлении различных очищающих смесей, таких как появление раствораХойера 26, внутренние структуры яйцеклетки ячменя были изображены с высокой степенью прозрачности, хотя ее эндосперм составляет основную часть семян. Эмбриогенез сахарной свеклы можно наблюдать путем очистки в сочетании с вакуумной обработкой и размягчения соляной кислотой19. Тем не менее, в отличие от видов, упомянутых выше, об эмбриологических наблюдениях путем очистки протоколов в семенах томатов не сообщалось. Это препятствует детальному исследованию эмбрионального и семенного развития томатов.
Хлоралгидрат обычно используется в качестве очищающего раствора, который позволяет погружаться тканям и клеткам отображаться на различных оптических плоскостях и по существу сохраняет клетки или тканевые компоненты 27,28,29. Протокол очистки на основе хлоралгидрата был успешно использован для очистки семян по всей установке для наблюдения за зародышем и эндоспермом Arabidopsis 21,28. Однако этот очистительный раствор не эффективен при очистке семян томатов, которые более непроницаемы, чем семена арабидопсиса. Физические барьеры включают: (1) кожный покров томата имеет почти 20 клеточных слоев при 3-15 DAF30,31, (2) эндосперм томата клеточного типа, а не ядерного типа32, и (3) семена томатов примерно в 70 раз больше поразмеру 33,34 и (4) производят большое количество слизи семенной оболочки, что блокирует проникновение очищающих реагентов и влияет на визуализацию эмбриональных клеток.
Таким образом, в настоящем отчете представлен оптимизированный метод очистки на основе хлоралгидрата для полной очистки семян томатов на разных стадиях, который позволяет глубоко визуализировать процесс развития эмбриона (рисунок 2).
