O tomate (S. lycopersicum L.) é uma das culturas vegetais mais importantes do mundo, com uma produção de 186,8 milhões de toneladas de frutos carnudos de 5,1 milhões de hectares em 20201. Pertence à grande família Solanaceae com cerca de 2.716 espécies2, incluindo muitas culturas comercialmente importantes, como berinjela, pimenta, batata e tabaco. O tomateiro cultivado é uma espécie diploide (2n = 2x = 24) com um tamanho de genoma de aproximadamente 900 Mb3. Durante muito tempo, um grande esforço tem sido feito para a domesticação e reprodução do tomate, selecionando características desejáveis de Solanum spp selvagem. Existem mais de 5.000 acessos de tomate listados no Tomato Genetics Resource Center e mais de 80.000 germoplasma de tomates são armazenados em todo o mundo4. A planta de tomate é perene na estufa e se propaga por sementes. Uma semente de tomate madura consiste em três compartimentos principais: um embrião adulto, endosperma residual do tipo celular e um revestimento de sementes duras 5,6 (Figura 1A). Após a dupla fertilização, o desenvolvimento de endospermas do tipo celular precede o desenvolvimento de zigotos. Em ~5-6 dias após a floração (DAF), o proembrião de duas células é observado pela primeira vez quando o endosperma consiste de seis a oito núcleos7. Em Solanum pimpinellifolium, o embrião se aproxima de seu tamanho final após 20 DAF, e as sementes são viáveis para germinação após 32 DAF8. À medida que o embrião se desenvolve, o endosperma é gradualmente absorvido e apenas uma pequena quantidade de endosperma permanece na semente. O endosperma residual consiste em endosperma micropilar ao redor da ponta da radícula e endosperma lateral no restante da semente 9,10. O revestimento externo da semente é desenvolvido a partir da epiderme externa espessada e lignificada do tegumento e, com as camadas mortas de restos de tegumento, formam uma casca dura para proteger o embrião e o endosperma5.
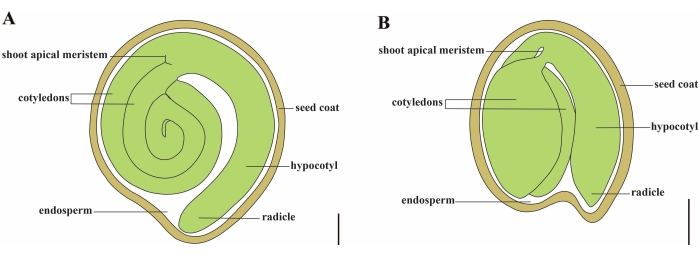
Figura 1: Representação esquemática de uma semente madura em Solanum lycopersicum e Arabidopsis thaliana. (A) Anatomia longitudinal de uma semente de tomateiro madura. (B) Anatomia longitudinal de uma semente madura de Arabidopsis. Uma semente de tomate madura é aproximadamente 70 vezes maior em tamanho do que uma semente de Arabidopsis. Barras de escala = (A) 400 μm, (B) 100 μm. Clique aqui para ver uma versão maior desta figura.
A produção de sementes de tomate de alta qualidade depende da coordenação entre o embrião, o endosperma e os componentes maternos das sementes11. Dissecar genes-chave e redes no desenvolvimento de sementes requer um registro fenotípico profundo e completo de sementes mutantes. Técnicas convencionais de embutimento-seccionamento, como a seção semifina e a seção de parafina, são amplamente aplicadas às sementes de tomateiro para observar as estruturas locais e mais finas do embrião12,13,14,15. No entanto, a análise do desenvolvimento de sementes a partir de seções finas é geralmente trabalhosa e carece de resolução espacial do eixo z. Em comparação, a depuração tecidual é um método rápido e eficiente para identificar o estágio de desenvolvimento dos defeitos embrionários com maior probabilidade de ocorrer16. O método de clareamento reduz a opacidade do tecido interno homogeneizando o índice de refração com um ou mais agentes bioquímicos16. A limpeza de tecidos inteiros permite a observação de uma estrutura tecidual vegetal sem destruir sua integridade, e a combinação de tecnologia de clareamento e imagens tridimensionais tornou-se uma solução ideal para obter informações sobre a morfologia e o estado de desenvolvimento de um órgão vegetal17,18. Ao longo dos anos, técnicas de limpeza de sementes têm sido utilizadas em várias espécies vegetais, incluindo Arabidopsis thaliana, Hordeum vulgare e Beta vulgaris19,20,21,22,23. Dentre estas, a tecnologia de limpeza de óvulos de montagem completa tem sido uma abordagem eficiente para o estudo do desenvolvimento de sementes de Arabidopsis, devido ao seu pequeno tamanho, 4-5 camadas da célula do revestimento da semente e o endosperma do tipo nuclear24,25. Com a atualização contínua de diferentes misturas de clareamento, como o surgimento da solução de Hoyer26, as estruturas internas do óvulo de cevada foram fotografadas com alto grau de clareza, embora seu endosperma constitua a maior parte das sementes. A embriogênese da beterraba sacarina pode ser observada por limpeza combinada com tratamento a vácuo e amolecimento com ácido clorídrico19. No entanto, ao contrário das espécies mencionadas acima, observações embriológicas por protocolos de limpeza em sementes de tomate não foram relatadas. Isso evita uma investigação detalhada sobre o desenvolvimento embrionário e de sementes de tomates.
O hidrato de cloral é comumente utilizado como solução de clareamento que permite que os tecidos e células imersos sejam exibidos em diferentes planos ópticos, preserva substancialmente as células ou componentes teciduais27,28,29. O protocolo de clareamento à base de hidrato de cloral tem sido utilizado com sucesso para a limpeza completa de sementes para observar o embrião e o endosperma de Arabidopsis21,28. No entanto, esta solução de limpeza não é eficiente na limpeza de sementes de tomate, que são mais impermeáveis do que as sementes de Arabidopsis. As barreiras físicas incluem: (1) o tegumento do tomate tem quase 20 camadas celulares em 3 a 15 DAF 30,31, (2) o endosperma do tomate é do tipo celular, não do tipo nuclear 32, e (3) as sementes de tomate são cerca de 70 vezes maiores em tamanho33,34 e (4) produzem grandes quantidades de mucilagem do revestimento da semente, o que bloqueia a penetração de reagentes de limpeza e afeta a visualização das células embrionárias.
Portanto, este relato apresenta um método otimizado de clareamento à base de hidrato de cloral para limpeza integral de sementes de tomateiro em diferentes estágios, o que permite imagens profundas do processo de desenvolvimento embrionário (Figura 2).
