El tomate (S. lycopersicum L.) es uno de los cultivos hortícolas más importantes del mundo, con una producción de 186,8 millones de toneladas de frutos carnosos de 5,1 millones de hectáreas en 20201. Pertenece a la gran familia Solanaceae con alrededor de 2.716 especies2, incluyendo muchos cultivos comercialmente importantes como berenjenas, pimientos, patatas y tabaco. El tomate cultivado es una especie diploide (2n = 2x = 24) con un tamaño de genoma de aproximadamente 900 Mb3. Durante mucho tiempo, se ha hecho un gran esfuerzo hacia la domesticación y reproducción del tomate mediante la selección de rasgos deseables de Solanum spp. silvestres. Hay más de 5.000 accesiones de tomate listadas en el Centro de Recursos Genéticos del Tomate y más de 80.000 germoplasma de tomates se almacenan en todo el mundo4. La planta de tomate es perenne en el invernadero y se propaga por semillas. Una semilla de tomate madura consta de tres compartimentos principales: un embrión adulto, endospermo residual de tipo celular y una capa dura de semilla 5,6 (Figura 1A). Después de la doble fertilización, el desarrollo del endospermo de tipo celular precede al desarrollo de cigotos. A ~ 5-6 días después de la floración (DAF), el proembrión bicelular se observa por primera vez cuando el endospermo consta de seis a ocho núcleos7. En Solanum pimpinellifolium, el embrión se acerca a su tamaño final después de 20 DAF, y las semillas son viables para la germinación después de 32 DAF8. A medida que el embrión se desarrolla, el endospermo se absorbe gradualmente y solo queda una pequeña cantidad de endospermo en la semilla. El endospermo residual consiste en endospermo micropilar que rodea la punta de la radícula, y endospermo lateral en el resto de la semilla 9,10. La capa externa de la semilla se desarrolla a partir de la epidermis externa engrosada y lignificada del tegumento, y con las capas muertas de restos de tegumento, forman una cáscara dura para proteger el embrión y el endospermo5.
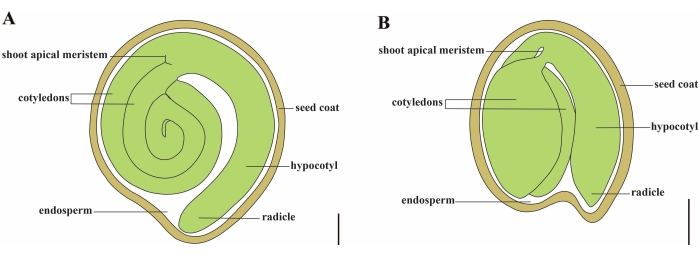
Figura 1: Representación esquemática de una semilla madura en Solanum lycopersicum y Arabidopsis thaliana. (A) Anatomía longitudinal de una semilla de tomate maduro. (B) Anatomía longitudinal de una semilla madura de Arabidopsis. Una semilla de tomate maduro es aproximadamente 70 veces más grande en tamaño que una semilla de Arabidopsis. Barras de escala = (A) 400 μm, (B) 100 μm. Haga clic aquí para ver una versión más grande de esta figura.
La producción de semillas de tomate de alta calidad depende de la coordinación entre el embrión, el endospermo y los componentes de la semilla materna11. La disección de genes y redes clave en el desarrollo de semillas requiere un registro fenotípico profundo y completo de semillas mutantes. Las técnicas convencionales de embutido-seccionamiento, como la sección semifina y la sección de parafina, se aplican ampliamente a las semillas de tomate para observar las estructuras locales y más finas del embrión12,13,14,15. Sin embargo, analizar el desarrollo de la semilla a partir de secciones delgadas suele ser laborioso y carece de resolución espacial del eje z. En comparación, la limpieza del tejido es un método rápido y eficiente para identificar la etapa de desarrollo de los defectos embrionarios que tienen más probabilidades de ocurrir16. El método de depuración reduce la opacidad del tejido interno homogeneizando el índice de refracción con uno o más agentes bioquímicos16. El aclaramiento de tejido completo permite observar la estructura de un tejido vegetal sin destruir su integridad, y la combinación de la tecnología de limpieza y la imagen tridimensional se ha convertido en una solución ideal para obtener información sobre la morfología y el estado de desarrollo de un órgano vegetal17,18. A lo largo de los años, las técnicas de limpieza de semillas se han utilizado en varias especies de plantas, incluyendo Arabidopsis thaliana, Hordeum vulgare y Beta vulgaris 19,20,21,22,23. Entre estos, la tecnología de limpieza de óvulos de montaje completo ha sido un enfoque eficiente para estudiar el desarrollo de semillas de Arabidopsis, debido a su pequeño tamaño, 4-5 capas de la célula de recubrimiento de la semilla y el endospermo de tipo nuclear24,25. Con la actualización continua de diferentes mezclas de aclaramiento, como la aparición de la solución de Hoyer26, las estructuras internas del óvulo de cebada fueron fotografiadas con un alto grado de claridad, aunque su endospermo constituye la mayor parte de las semillas. La embriogénesis de la remolacha azucarera se puede observar mediante la limpieza combinada con el tratamiento al vacío y el ablandamiento con ácido clorhídrico19. Sin embargo, a diferencia de las especies mencionadas anteriormente, no se han reportado observaciones embriológicas mediante protocolos de limpieza en semillas de tomate. Esto impide una investigación detallada sobre el desarrollo embrionario y de semillas de los tomates.
El hidrato de cloral se utiliza comúnmente como una solución de limpieza que permite que los tejidos y células sumergidos se muestren en diferentes planos ópticos, y preserva sustancialmente las células o componentes tisulares27,28,29. El protocolo de limpieza basado en hidrato de cloral se ha utilizado con éxito para el aclaramiento de semillas de montaje completo para observar el embrión y el endospermo de Arabidopsis21,28. Sin embargo, esta solución de limpieza no es eficiente para limpiar las semillas de tomate, que son más impermeables que las semillas de Arabidopsis. Las barreras físicas incluyen: (1) el tegumento del tomate tiene casi 20 capas celulares de 3 a 15 DAF 30,31, (2) el endospermo del tomate es de tipo celular, no de tipo nuclear 32, y (3) las semillas de tomate son aproximadamente 70 veces más grandes en tamaño33,34 y (4) producen grandes cantidades de mucílago de recubrimiento de semillas, lo que bloquea la penetración de los reactivos de limpieza y afecta la visualización de las células embrionarias.
Por lo tanto, este informe presenta un método optimizado de limpieza basado en hidrato de cloral para la limpieza de semillas de tomate de montaje completo en diferentes etapas, lo que permite obtener imágenes profundas del proceso de desarrollo embrionario (Figura 2).
