टमाटर (एस लाइकोपर्सिकम एल) दुनिया भर में सबसे महत्वपूर्ण सब्जी फसलों में से एक है, जिसमें 2020 में 5.1 मिलियन हेक्टेयर से 186.8 मिलियन टन मांसल फलों का उत्पादन होता है। यह लगभग 2,716 प्रजातियों2 के साथ बड़े सोलानासी परिवार से संबंधित है, जिसमें बैंगन, मिर्च, आलू और तंबाकू जैसी कई व्यावसायिक रूप से महत्वपूर्ण फसलें शामिल हैं। खेती की गई टमाटर एक द्विगुणित प्रजाति (2n = 2x = 24) है जिसका जीनोम आकार लगभग 900 Mb3 है। लंबे समय से, जंगली सोलनम एसपीपी से वांछनीय लक्षणों का चयन करके टमाटर को पालतू बनाने और प्रजनन की दिशा में बहुत प्रयास किया गया है। टमाटर जेनेटिक्स रिसोर्स सेंटर में 5,000 से अधिक टमाटर परिग्रहण सूचीबद्ध हैंऔर टमाटर के 80,000 से अधिक जर्मप्लाज्म दुनिया भर में संग्रहीत हैं। टमाटर का पौधा ग्रीनहाउस में बारहमासी होता है और बीजों द्वारा फैलता है। एक परिपक्व टमाटर के बीज में तीन प्रमुख डिब्बे होते हैं: एक पूर्ण विकसित भ्रूण, अवशिष्ट सेलुलर-प्रकार एंडोस्पर्म, और एक कठोर बीज कोट 5,6 (चित्रा 1 ए)। दोहरे निषेचन के बाद, सेलुलर-प्रकार के एंडोस्पर्म का विकास युग्मनज के विकास से पहले होता है। फूल (डीएएफ) के ~ 5-6 दिनों के बाद, दो-कोशिका वाले प्रोम्ब्रायो को पहली बार देखा जाता है जब एंडोस्पर्म में छह से आठ नाभिकहोते हैं। सोलनम पिंपिनेलिफोलियम में, भ्रूण 20 डीएएफ के बाद अपने अंतिम आकार तक पहुंचता है, और बीज 32 डीएएफ 8 के बाद अंकुरण के लिए व्यवहार्यहोते हैं। जैसे-जैसे भ्रूण विकसित होता है, एंडोस्पर्म धीरे-धीरे अवशोषित हो जाता है और बीज में एंडोस्पर्म की केवल थोड़ी मात्रा रह जाती है। अवशिष्ट एंडोस्पर्म में रेडियल टिप के आसपास माइक्रोपाइलर एंडोस्पर्म और शेष बीज 9,10 में पार्श्व एंडोस्पर्म होते हैं। बाहरी बीज कोट को इन्टेगुमेंट के मोटे और लिग्निफाइड बाहरी एपिडर्मिस से विकसित किया जाता है, और इन्टेगुमेंट अवशेषों की मृत परतों के साथ, वे भ्रूण और एंडोस्पर्म5 की रक्षा के लिए एक कठोर खोल बनाते हैं।
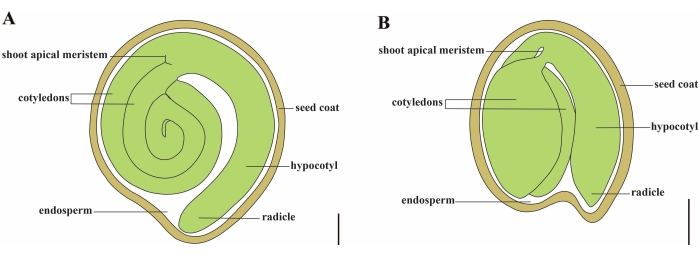
चित्र 1: सोलनम लाइकोपर्सिकम और एराबिडोप्सिस थैलियाना में एक परिपक्व बीज का योजनाबद्ध प्रतिनिधित्व। (ए) एक परिपक्व टमाटर के बीज की अनुदैर्ध्य शारीरिक रचना। (बी) एक परिपक्व एराबिडोप्सिस बीज की अनुदैर्ध्य शारीरिक रचना। एक परिपक्व टमाटर का बीज एराबिडोप्सिस बीज की तुलना में आकार में लगभग 70 गुना बड़ा होता है। स्केल पट्टियाँ = (A) 400 μm, (B) 100 μm. कृपया इस आंकड़े का एक बड़ा संस्करण देखने के लिए यहां क्लिक करें.
उच्च गुणवत्ता वाले टमाटर के बीज का उत्पादन भ्रूण, एंडोस्पर्म और मातृ बीज घटकों के बीच समन्वय पर निर्भर करताहै। बीज विकास में प्रमुख जीन और नेटवर्क को विच्छेदित करने के लिए उत्परिवर्ती बीजों की गहरी और पूर्ण-ट्रैक फेनोटाइपिक रिकॉर्डिंग की आवश्यकता होती है। पारंपरिक एम्बेडिंग-सेक्शनिंग तकनीक, जैसे अर्ध-पतले खंड और पैराफिन अनुभाग, भ्रूण 12,13,14,15 की स्थानीय और महीन संरचनाओं का निरीक्षण करने के लिए टमाटर के बीज पर व्यापक रूप से लागू होते हैं। हालांकि, पतले वर्गों से बीज विकास का विश्लेषण करना आमतौर पर श्रमसाध्य होता है और जेड-अक्ष स्थानिक संकल्प का अभाव होता है। इसकी तुलना में, ऊतक समाशोधन भ्रूण दोषों के विकास चरण को इंगित करने के लिए एक तेज़ और कुशल तरीका हैजो होने की सबसे अधिक संभावना है। समाशोधन विधि एक या अधिक जैव रासायनिक एजेंटों के साथ अपवर्तक सूचकांक को समरूप करके आंतरिक ऊतक की अपारदर्शिता को कम करतीहै। संपूर्ण ऊतक समाशोधन इसकी अखंडता को नष्ट किए बिना एक पौधे के ऊतक संरचना के अवलोकन की अनुमति देता है, और समाशोधन तकनीक और त्रि-आयामी इमेजिंग का संयोजन एक पौधे के अंग17,18 की आकृति विज्ञान और विकासात्मक स्थिति पर जानकारी प्राप्त करने के लिए एक आदर्श समाधान बन गया है। वर्षों से, विभिन्न पौधों की प्रजातियों में बीज समाशोधन तकनीकों का उपयोग किया गया है, जिसमें एराबिडोप्सिस थैलियाना, होर्डियम वल्गर और बीटा वल्गरिस19,20,21,22,23 शामिल हैं। इनमें से, होल-माउंट डिंब समाशोधन तकनीक एराबिडोप्सिस के बीज विकास का अध्ययन करने के लिए एक कुशल दृष्टिकोण रही है, इसके छोटे आकार, बीज कोट सेल की 4-5 परतों और परमाणु-प्रकार एंडोस्पर्म24,25 के कारण। विभिन्न समाशोधन मिश्रणों के निरंतर अद्यतन के साथ, जैसे कि होयर के समाधान26 के उद्भव के साथ, जौ के अंडाणु की आंतरिक संरचनाओं को उच्च स्तर की स्पष्टता के साथ चित्रित किया गया था, हालांकि इसके एंडोस्पर्म बीज का बड़ा हिस्सा बनाते हैं। चीनी बीट के भ्रूणजनन को वैक्यूम उपचार के साथ संयुक्त रूप से साफ़ करके और हाइड्रोक्लोरिक एसिड19 के साथ नरम करके देखा जा सकता है। बहरहाल, ऊपर उल्लिखित प्रजातियों के विपरीत, टमाटर के बीज में प्रोटोकॉल को साफ़ करके भ्रूण संबंधी अवलोकनों की सूचना नहीं दी गई है। यह टमाटर के भ्रूण और बीज विकास में विस्तृत जांच को रोकता है।
क्लोरल हाइड्रेट का उपयोग आमतौर पर एक समाशोधन समाधान के रूप में किया जाता है जो डूबे हुए ऊतकों और कोशिकाओं को विभिन्न ऑप्टिकल विमानों पर प्रदर्शित करने की अनुमति देता है, और कोशिकाओं या ऊतक घटकों 27,28,29 को काफी हद तक संरक्षित करता है। एराबिडोप्सिस21,28 के भ्रूण और एंडोस्पर्म का निरीक्षण करने के लिए बीजों के पूरे माउंट क्लियरिंग के लिए क्लोरल हाइड्रेट-आधारित क्लियरिंग प्रोटोकॉल का सफलतापूर्वक उपयोग किया गया है। हालांकि, यह समाशोधन समाधान टमाटर के बीज को साफ करने में कुशल नहीं है, जो एराबिडोप्सिस बीजों की तुलना में अधिक अभेद्य हैं। भौतिक बाधाओं में शामिल हैं: (1) टमाटर के आंत में 3 से 15 डीएएफ 30,31 पर लगभग20 सेल परतें होती हैं, (2) टमाटर एंडोस्पर्म सेलुलर-प्रकार होता है, परमाणु-प्रकार32 नहीं, और (3) टमाटर के बीज आकार में लगभग 70 गुना बड़ेहोते हैं 33,34 और (4) बड़ी मात्रा में बीज कोट म्यूसिलेज का उत्पादन करते हैं, जो समाशोधन अभिकर्मकों के प्रवेश को अवरुद्ध करता है और भ्रूण कोशिकाओं के दृश्य को प्रभावित करता है।
इसलिए, यह रिपोर्ट विभिन्न चरणों में टमाटर के बीज के पूरे माउंट क्लियरिंग के लिए एक अनुकूलित क्लोरल हाइड्रेट-आधारित समाशोधन विधि प्रस्तुत करती है, जो भ्रूण विकास प्रक्रिया की गहरी इमेजिंग की अनुमति देती है (चित्रा 2)।
