Gezien de prevalentie van slaap in onze dagelijkse routine, is het belangrijk om zijn functie te begrijpen. Studies met deze doelstelling vereisen een precieze meting van de slaap. Polysomnography (PSG) is de goud-standaard maat van de slaap. PSG zorgt voor objectieve, kwantitatieve meting van de slaap met een hoge temporele resolutie en kan nuttig zijn voor zowel onderzoek als klinische doeleinden. PSG is een combinatie van fysiologische opnames. Een PSG-montage omvat minimaal de volgende maatregelen: elektro-encefalografie (EEG), elektrooculografie (EOG) en elektro myografie (EMG). Deze maatregelen beoordelen elektrische potentialen van de hersenen, de ogen, en spieren respectievelijk, en toestaan voor de indeling van de slaap stadia (Zie Figuur 1). Andere maatregelen, zoals elektrocardiografie (ECG), ademhaling, en Pulse oximetrie kunnen worden opgenomen om de aanwezigheid van ongeordende slaap te identificeren.
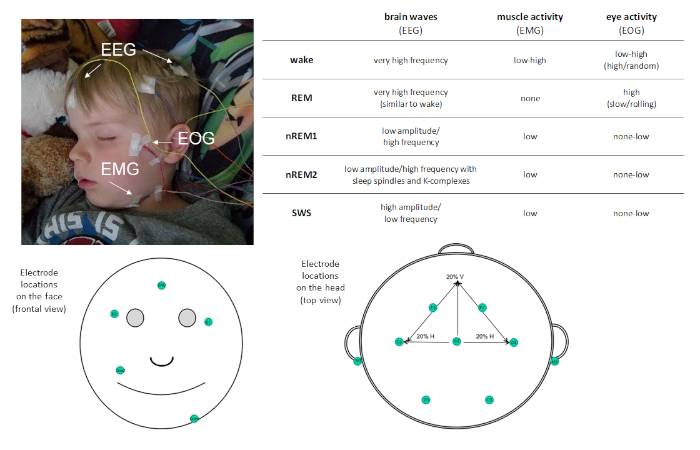
Figuur 1: voorbeeld van de plaatsing van de elektrode en beschrijving van de activiteit vastgelegd via PSG. Klik hier om een grotere versie van dit cijfer te bekijken.
PSG laat slaap worden gekarakteriseerd in vier verschillende slaap stadia: niet-snelle oogbeweging (Non-REM) fase 1 (nREM1; 4 − 7 Hz), Non-REM fase 2 (nREM2; 12 − 15 Hz) en Non-REM fase 3 (beter bekend als Slow Wave Sleep [SWS]; 0,5 − 4 Hz) en Rapid-Eye-beweging (REM slaap. nREM1 markeert de slaap begin, en wordt geïdentificeerd op basis van verminderde spierspanning in de EMG heroverd en gemengde amplitude EEG oscillaties ten opzichte van de Alfa waargenomen in rust wakker. Dit wordt gevolgd door nREM2, die kan worden onderscheiden door de aanwezigheid van slaap spindels (korte uitbarstingen van Sigma frequentie activiteit; 11 − 16 Hz) en K-complexen (enkele langzame golven die zich onderscheiden van de omringende activiteit) in het EEG. SWS wordt gekenmerkt door verschillende langzame frequentie-oscillaties met hoge amplitude. REM-slaap wordt gekenmerkt door snelle lage-amplitude oscillerende hersenactiviteit zeer vergelijkbaar met die waargenomen tijdens de Wake. Echter, wat REM-slaap van wakker onderscheidt, is dat het ook wordt gekenmerkt door fasische snelle oogbewegingen (vandaar de moniker REM) en spier atonia. In de loop van een slaap bout, slaap stadia worden cyclisch ervaren, met een snelheid van ongeveer 90 min/cyclus.
Slaap volgt ook het circadiane ritme, met slaap aanvallen plaatsvinden in 24-h cycli. Slaap timing en consistentie kunnen invloed hebben op de slaapfunctie en zijn ook belangrijk om te beoordelen. Hoewel PSG noodzakelijk is om slaap stadia te karakteriseren, is het tijdrovend om toe te passen en daarom niet ideaal voor het beoordelen van meerdere slaap aanvallen (bijv. meerdere nachten slaap, nap’s en nachtrust). Voor dit, actigrafie is gunstig. Actigrafie maakt gebruik van een Tri-axiale versnellingsmeter, meestal op de pols, om de slaap te schatten op basis van de afwezigheid van beweging. Hoewel actigrafie niet kan worden gebruikt om de slaap stadia te karakteriseren, is aangetoond dat het betrouwbaar is bij het detecteren van de slaap begin en het begin van de Wake (inclusief slaap fragmentatie of ontwaken na slaap begin) in een reeks populaties van baby’s1 tot oudere volwassenen2 . Zowel PSG als actigrafie zijn de voorkeurs methoden voor zelf-/moederrapport metingen. Zelf/ouder-rapport maatregelen zijn eenvoudig te beheren en relatief goedkoop, maar ze zijn ook onderhevig aan bias en niet-naleving. Ten slotte is het vermeldenswaard dat deze methoden kunnen worden gebruikt in combinatie om te kapitaliseren op de sterke punten van elk. Zo kan PSG worden gecombineerd met actigrafie en/of zelf/ouder-rapport om zowel de nachtelijke slaapkwaliteit te verkrijgen als de controle van slaap hoeveelheden of slaap-waak cycli, vooral over lange duur (bijv. weken of maanden).
Een functie van de slaap die bijzondere interesse heeft vergaard is slaap-afhankelijke geheugen consolidatie, de verwerking van herinneringen die hen sterker en minder kwetsbaar voor interferentie3laat. Hoewel geheugen consolidatie kan plaatsvinden tijdens de wake bij kinderen4 en volwassenen5, is er aanzienlijke bewijs dat consolidatie wordt verbeterd tijdens de slaap. Afgelopen onderzoek heeft gedrags voorbeelden van slaap-afhankelijke geheugen consolidatie gegeven door het vergelijken van veranderingen in de geheugenprestaties na een slaap interval (bijv. 8 pm − 8 am) tot veranderingen na een gelijkwaardig interval dat wakker is (bijv. 8 am − 8 pm). Bij volwassenen, herinneringen worden beschermd6 of zelfs verbeterd7 na een slaap interval terwijl herinneringen meestal verval over een equivalente interval van Wake. Er zijn besturingselementen gebruikt die de prestatie wijzigingen van circadiane invloeden op8,9,10dissociëren. Vergelijkbare voordelen van slaap worden bijvoorbeeld waargenomen bij het vergelijken van prestaties over een Mid-Day NAP naar een equivalente Mid-Day Wake period9.
Hoewel slaap ooit werd gedacht aan een passief proces weerspiegelen, gewoon het beschermen van herinneringen van verval of interferentie, moderne theorieën suggereren slaap speelt een actievere rol en eigenlijk bevordert geheugen door heractiveringen11,12 ,13. Ondersteuning voor dit komt uit waargenomen correlaties tussen gedragsmaatregelen van geheugen consolidatie over de slaap (verandering in geheugen terugroepen na slaap in vergelijking met voor slaap) en specifieke aspecten van slaap fysiologie. Voor veel declaratieve geheugen taken, geheugen consolidatie wordt geassocieerd met aspecten van non-REM-slaap, specifiek maatregelen van SWS of slaap spindels gevonden in nREM2 en SWS. Als de rol van de slaap passieve bescherming tegen interferentie was, zou een dergelijke correlatie niet worden verwacht; eerder een correlatie tussen de tijd in slaap (ongeacht de slaap fase) en de prestaties zou worden verwacht, als meer tijd in slaap zou bieden meer bescherming tegen interferentie14.
Aanvullende ondersteuning voor de actieve rol van SWS in geheugen consolidatie is duidelijk in studies van gerichte geheugen reactivering. In deze studies, een geheugen wordt geleerd in de context van een perceptuele Hint, bijvoorbeeld een geur, en terugroepen van het geheugen is groter na de slaap als de hint opnieuw wordt gepresenteerd tijdens de slaap, SWS in het bijzonder15. Hoewel het onderliggende mechanisme wordt besproken16,17, een prominente theorie, systemen consolidatie theorie, beweert dat herinneringen gecodeerd in de Hippocampus worden gestabiliseerd in de cortex hoewel hippocampal-neocortical dialoog. Specifiek, corticale langzame golven en slaap spindels, die optreden in combinatie met hippocampal rimpelingen geassocieerd met geheugen reactivering, ondersteuning van de geheugen overdracht3.
De rol van slaap in geheugen consolidatie tijdens de ontwikkeling is minder duidelijk. Vroege kindertijd is een periode van bijzonder belang als kinderen beginnen te overstappen van een bifasische (bestaande uit een dutje van een dag en een nachtelijke slaap bout) naar een monofasisch slaappatroon. Recent onderzoek suggereert dat deze overgang kan weerspiegelen de hersenen rijping18. Dit argument is consistent met empirische gegevens die veranderingen in de ontwikkeling van de nachtrust (d.w.z. topografie van langzame Golf activiteit) spiegelen die van corticale rijping19.
Hoewel er verschillende gedrags demonstraties van de nachtelijke slaap-afhankelijke consolidatie bij kinderen20,21 en zuigelingen22, onderzoek naar de neurale onderbouwing van geheugen consolidatie met Mid-Day slaap zijn net beginnen te worden onderzocht. In baanbrekend werk dat de rol van Mid-Day nap’s op het geheugen in voorschoolse kinderen onderzoekt, werden nap’s getoond om herinneringen van recent geleerde informatie te beschermen, terwijl het geheugen werd verlaagd (door ~ 12%) Wanneer kinderen wakker bleven tijdens het NAP-interval23. Dit “NAP-voordeel” was het grootst bij kinderen die gewoonlijk (d.w.z. 5 of meer keer per week, zoals gemeten met actigrafie), onafhankelijk van hun leeftijd, hebben aangeboord. Door het opnemen van PSG tijdens het NAP bleek de verandering in de geheugenprestaties over de NAP-periode specifiek te zijn gekoppeld aan de slaap-spil dichtheid (het aantal slaap spindels per minuut van nREM), wat suggereert dat NAP-kwaliteit (geen hoeveelheid) een kritieke factor was in bevordering van geheugen retentie (Zie de sectie representatieve resultaten).
Deze studie belicht de betekenis van PSG in het verkennen van de relaties tussen slaap en geheugen tijdens de ontwikkeling. Het wijst op het belang van het karakteriseren van slaap macro-(slaap stadia) en micro-(kwaliteiten van die stadia zoals frequenties en de aanwezigheid van spindels) structuren tijdens nap’s voor geheugen consolidatie. Het benadrukt ook het belang van het beoordelen van slaap ritmes (karakteriseren van kinderen als gewone of niet-gewone nappers). Hoewel ons werk de functie van nap’s in visuospatial learning heeft gekarakteriseerd (en meer recentelijk emotionele24 en procedurele25 -leren), blijven er veel vragen over. Het is bijvoorbeeld belangrijk om andere declaratieve geheugen taken te onderzoeken om de generalizability van deze bevindingen te beoordelen en om taken te beoordelen die in voorschoolse klaslokalen worden gebruikt om specifieke parameters te begrijpen (bijvoorbeeld het bedrag van NAP-voordeel ten opzichte van leren) voor ecologisch geldige taken. Extra werk zal ook nodig zijn om te begrijpen wanneer wakker voldoende is voor geheugen consolidatie. Ons doel is dus om het proces van het meten van slaap en slaap-afhankelijke geheugen consolidatie bij kinderen te demystificeren. We bieden praktische tips voor het onderzoeken van het voordeel van een middagdutje op declaratief geheugen bij het ontwikkelen van kleuters (ongeveer 3 tot 4 jaar oud) met behulp van een computergestuurde visuospatiale geheugen taak en methoden voor het beoordelen van gegemanierd NAP met behulp van actigrafie, ouder-rapport en NAP-fysiologie met behulp van PSG. Hoewel deze methoden zijn ontwikkeld voor voorschoolse leeftijd kinderen die met wisselende frequentie NAP, deze methoden kunnen worden aangepast aan elke leeftijdsgroep.
