השכיחות של השינה בשגרה היומית שלנו, חשוב להבין את תפקידה. מחקרים עם מטרה זו דורשים מדידה מדויקת של שינה. פוליסומגרפיה (PSG) הוא מדד בתקן זהב של שינה. PSG מאפשר אובייקטיבי, מדידה כמותית של שינה עם רזולוציה גבוהה בזמן והוא יכול להיות שימושי הן מחקר ומטרות קליניות. PSG הוא שילוב של הקלטות פיזיולוגיות. לפחות, מונטאז א כולל את הצעדים הבאים: אלקטרונצגרפיה (EEG), אלקטרונוקולוגרפיה (EOG), ואלקטרומגרפיה (EMG). אמצעים אלה משלבים את הפוטנציאל החשמלי מהמוח, העיניים והשרירים בהתאמה, ומאפשרים סיווג של שלבי שינה (ראה איור 1). אמצעים אחרים, כגון אלקטרוקרדיוגרפיה (א. ג. א), נשימה וחמצן עשויים להיכלל כדי לזהות נוכחות של שינה מלא מעשית.
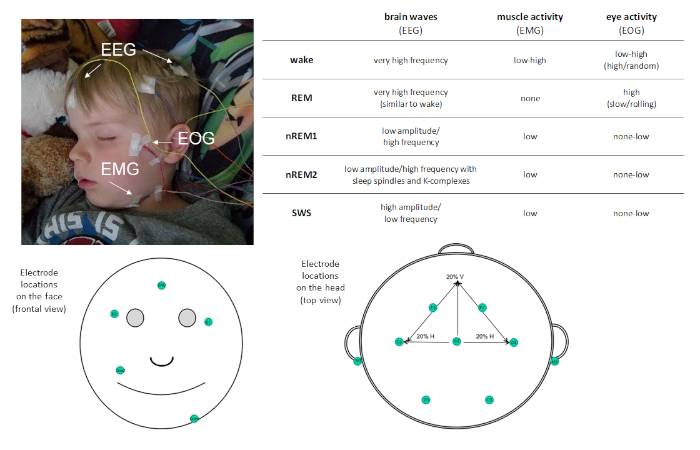
איור 1: מיקום האלקטרודה לדוגמה ותיאור הפעילות שנרשמה באמצעות PSG. אנא לחץ כאן כדי להציג גירסה גדולה יותר של איור זה.
PSG מאפשר לישון להיות מאופיין לארבעה שלבי שינה נפרדים: תנועת עיניים שאינה מהירה (שאינה REM) שלב 1 (nREM1; 4-7 Hz), לא-ראם שלב 2 (nREM2; 12-15 Hz), ושלא-ראם שלב 3 (הידוע יותר בכינויו שינת גל איטית [SWS]; 0.5-4 Hz) ותנועת עיניים מהירה (REM ישון. nREM1 מציין התפרצות שינה, והוא מזוהה על בסיס טונוס שרירים מופחתת ב-EMG ההמרה ואת משרעת מעורבת EEG תנודות ביחס אלפא נצפתה בהתעוררות מנוחה. זה ואחריו nREM2, אשר יכול להיות מכובד על ידי נוכחות של הצירים שינה (התפרצויות קצרות של פעילות תדר סיגמא; 11-16 Hz) ו-K-מתחמי (בודד איטי גלים כי להתבלט מן הפעילות הסובבת) ב-EEG. SWS מאופיין על ידי שונות בתדר איטי באמצעות משרעת גבוהה EEG תנודות. שינה REM מאופיינת במהירות נמוכה משרעת פעילות המוח מנדנוד דומה מאוד לזה שנצפה במהלך ההתעוררות. עם זאת, מה המבדיל REM שינה מן ההתעוררות היא כי הוא מאופיין גם על ידי תנועות העין phasic מהירה (ומכאן REM כינוי) ושריר atonia. במהלך התחרות שינה, שלבי השינה מנוסים מחזוריות, בקצב של כ 90 דקות/מחזור.
שינה גם מלווה את קצב המעגל, עם התקפי שינה המתרחשים 24-h מחזורים. תזמון שינה ועקביות עשויים להשפיע על תפקוד השינה וחשוב גם להעריך. למרות PSG יש צורך לאפיין את שלבי השינה, זה זמן רב ליישם ולכן לא אידיאלי להערכת התקפי שינה מרובים (למשל, לילות מרובות של שינה, תנומות ושינה לילה). . בשביל זה, האקטגרפיה מועילה אקטגרפיה משתמשת בתאוצה tri-צירית, בדרך כלל על פרק כף היד, כדי להעריך את השינה על בסיס העדר תנועה. למרות האקטגרפיה לא ניתן להשתמש כדי לאפיין את שלבי השינה, זה הוכח להיות אמין בגילוי התפרצות שינה והתעוררות ההתעוררות (כולל פיצול שינה או להתעורר לאחר הופעת השינה) במגוון של אוכלוסיות מתינוקות1 למבוגרים2 . הן PSG ו האקטגרפיה הן שיטות מועדפות באמצעות מדדים עצמית/הורה-דוח. מדידות עצמית/הורה-דוח קל לנהל וזול יחסית, עם זאת, הם כפופים גם הטיה ואי ציות. לבסוף, ראוי לציין כי ניתן להשתמש בשיטות אלה בשילוב כדי לנצל את היתרונות של כל אחד מהם. לדוגמה, PSG יכול להיות משולב עם האקטגרפיה ו/או באמצעות דו ח עצמי/ההורה כדי לקבל הן איכות השינה לילה, כמו גם אימות של כמויות שינה או מחזורי התעוררות השינה, במיוחד על משכים ארוכים (למשל, שבועות או חודשים).
פונקציה אחת של שינה שצברה עניין מיוחד היא איחוד זיכרון תלויי שינה, עיבוד הזיכרונות שמשאיר אותם חזקים יותר ופגיעים פחות להפרעות3. למרות שאיחוד הזיכרון יכול להתקיים במהלך ההתעוררות אצל ילדים4 ומבוגרים5, יש ראיות משמעותיות כי האיחוד הוא משופר במהלך השינה. מחקרים שנערכו בעבר סיפקו דוגמאות התנהגותיות לאיחוד זיכרון תלוי-שינה על-ידי השוואת שינויים בביצועי זיכרון לאחר מרווח של שינה (לדוגמה, 08:08:00) לשינויים בעקבות מרווח שווה ערך שהושקע בערה (לדוגמה, 08:08:00). במבוגרים, זיכרונות מוגנים6 או אפילו משופר7 בעקבות מרווח של שינה בעוד הזיכרונות בדרך כלל להירקב על פני מרווח שווה ערך של התעוררות. מועסקים בפקדים שאינם משתמשים בשינויים בביצועים מהשפעות אחראי8,9,10. לדוגמה, יתרונות דומים של שינה נשמרים בעת השוואת ביצועים במהלך שינה באמצע היום ועד לתקופת התעוררות שוות-ערך של אמצע היום9.
למרות ששינה חשבה לשקף תהליך פסיבי, פשוט להגן על זיכרונות מפני ריקבון או הפרעה, תיאוריות מודרניות מרמזות על שינה תפקיד פעיל יותר ולמעשהמקדמת את הזיכרון באמצעותהפעלות 11,12 ,13. תמיכה עבור זה נובע מיחסי הציבור הנצפים בין צעדים התנהגותיים של קונסולידציה זיכרון על פני השינה (לשנות את הזיכרון להיזכר לאחר השינה לעומת לפני השינה) והיבטים ספציפיים של פיזיולוגיה שינה. עבור משימות זיכרון הצהרתי רבות, איחוד זיכרון משויך להיבטים של שינה שאינה REM, במיוחד מדדים של SWS או הצירים לשינה שנמצאו ב-nREM2 ו-SWS. אם תפקיד השינה היה הגנה פסיבית מפני הפרעות, מתאם כזה לא צפוי; מתאם בין הישנות הזמן (ללא קשר לשלב השינה) והביצועים צפויים, ככל ששינה רבה יותר תספק הגנה רבה יותר מהפרעות14.
תמיכה נוספת עבור התפקיד הפעיל של SWS באיחוד זיכרון ניכרת במחקרים של הפעלה מראש של זיכרון ממוקד. במחקרים אלה, הזיכרון נלמד בהקשר של הרמז התפיסתי, למשל ריח, והחזרה של הזיכרון הוא יותר לאחר השינה אם האות מוצג מחדש במהלך השינה, SWS בפרט15. למרות המנגנון הבסיסי הוא התווכחו16,17, אחת תיאוריה בולטת, המערכת התיאוריה קונסולידציה, לטענת כי הזיכרונות המקודדים בהיפוקמפוס מיוצב בקליפת המוח למרות היפוקמאל-הידברות דיאלוג. במיוחד, גלי המוח איטי והצירים לישון, המתרחשים בשילוב עם גלים היפוקמאל הקשורים ההפעלה זיכרון, תמיכה העברת זיכרון3.
תפקיד השינה באיחוד הזיכרון במהלך הפיתוח הוא פחות ברור. הילדות המוקדמת היא תקופה של עניין מיוחד כאשר הילדים מתחילים לעבור מתוך biphasic (המורכב של תנומה באמצע היום והתקף שינה לילה) לתבנית שינה מונוחסית. מחקרים אחרונים מראים כי מעבר זה עשוי לשקף התבגרות המוח18. טיעון זה מתאים לנתונים אמפיריים המציגים שינויים התפתחותיים בשינה לילה (כלומר, טופוגרפיה של פעילות גל איטית) מראות של התבגרות בקליפת הראש19.
למרות שישנן מספר הפגנות התנהגותיות של איחוד תלוי לילה בילדים20,21 ו תינוקות22, מחקר על התחתון נוירופינגס של איחוד זיכרון עם שינה באמצע היום הם רק מתחיל להיחקר. בעבודה פורצת דרך לבחון את התפקיד של התנומות באמצע היום על הזיכרון בגן הילדים, התנומות הוכחו כדי להגן על זיכרונות של מידע שנלמד לאחרונה, בעוד זיכרון הופחת (על ידי ~ 12%) כשהילדים נשארו ערים בזמן השינה23. זה “יתרון שינה” היה הגדול ביותר אצל ילדים שהיו מנמנמת (כלומר, 5 פעמים או יותר בשבוע כפי שנמדד עם האקטגרפיה) ללא קשר לגילם. על-ידי הקלטת psg במהלך התנומה, השינוי בביצועי הזיכרון לאורך תקופת התנומה נמצא קשור במיוחד לצפיפות ציר השינה (מספר הצירים לשינה לדקה של nrem), הרומז על איכות התנומה (לא כמות) היה גורם קריטי ב קידום שמירת הזיכרון (עיין בסעיף התוצאות המייצג).
מחקר זה מדגיש את המשמעות של PSG בחקר היחסים בין שינה לבין זיכרון במהלך הפיתוח. הוא מצביע על החשיבות של אפיון מאקרו שינה-(שלבי השינה) ומיקרו-(איכויות של שלבים כגון תדרים ונוכחות של צירים) מבנים במהלך התנומות לאיחוד זיכרון. כמו-כן, היא מדגישה את החשיבות של הערכת מקצבים שינה (אפיון הילדים כנאפה או שאינו הרגל הרגיל). למרות עבודתנו יש לאפיין את הפונקציה של תנומות בלמידה ויזואומרחבית (ולאחרונה רגשית24 ו פרוצדורליים25 לימוד), שאלות רבות נותרו. למשל, יהיה חשוב לבדוק משימות זיכרון הצהרתי אחרות כדי להעריך את היכולת המידיתית של ממצאים אלה ולהעריך משימות המשמשות כיתות גן כדי להבין פרמטרים ספציפיים (למשל, כמות הטבה לנמנום ביחס ללמידה) עבור משימות חוקיות מבחינה אקולוגית. עבודה נוספת תהיה נחוצה גם כדי להבין מתי ההתעוררות מספיקה לאיחוד הזיכרון. לפיכך, המטרה שלנו היא להכפיש את התהליך של מדידת שינה ושינה בזיכרון לקונסולידציה בילדים. אנו מספקים עצות מעשיות לבדיקת היתרון של נמנום אחר הצהריים על זיכרון הצהרתי בדרך כלל לפתח גיל הרך (כ 3 עד 4 שנים של גיל) באמצעות משימה ממוחשבת הזיכרון ויזואומרחבית כמו גם שיטות להערכת נמנום habituality באמצעות הקטיגרפיה, דוח הורים ופיזיולוגיה של nap באמצעות PSG. למרות ששיטות אלה פותחו עבור ילדים בגיל הרך התנומה בתדירות שונים, שיטות אלה יכול להיות מותאם לכל קבוצת גיל.
