The procedure of RB generation is elucidated in the Figure 1, which combines the adherent and floating culture. It was possible to harvest the human RB from RB1-KO hESC, and obtain the RB cell line by isolating the RB organoids.
Here, the protocol provides the details of the differentiation in different stages (Figure 2). Hollow spheres are formed in the first 3 days which attach to the culture surface and then expand (Figure 2A-E). From day 15 onward, cells are elevated and culture in suspension (Figure 2F). The day after the detachment, retinal organoids are formed, and the bright rims are visible (Figure 2G, black arrows). Moreover, those cells outside the organoids are likely to die in the following week (Figure 2G, orange arrows). On day 27, the optic vesicle architecture is evident and around 90% of organoids display this structure (Figure 2H); the organoids without this structure could be discarded. The first detection of the RB occurs on day 45, and then it becomes palpable on day 50 (Figure 2I). When it grows to day 90, the optic vesicle structures are principally enfolded by the RB (Figure 2J). Meanwhile, the RB could be isolated as an RB cell line for further culture (Figure 2K). Above 80% retinal organoids would be fully enveloped by the RB on day 105 (Figure 2L). They highly show expression of Ki67 (proliferation marker) and SYK (oncogene marker) comparing with the H9-derived retinal organoids, which indicates the tumorigenesis in the RB organoids (Figure 2M,N). Additionally, the high expression of ARR3 (cone precursor maker) and CRX (photoreceptor precursor marker) in the RB organoids demonstrates that they originate from cone precursor cells (Figure 2O,P).
The procedure of RB generation mainly undergoes three stages with morphology changes before the RB formation; here, the study provides the inferior and superior results at those stages (Figure 3). Differentiated and undifferentiated hESC (Figure 3A,B) is easy to distinguish from the morphology, and the undifferentiated hESC is chosen for RB formation. On day 5, a hollow sphere should generate (Figure 3D) rather than the solid one (Figure 3C). The RB is derived from the retinal organoids, which display optic vesicle architecture (Figure 3F). There is no RB that would generate in the inferior organoids (Figure 3E).
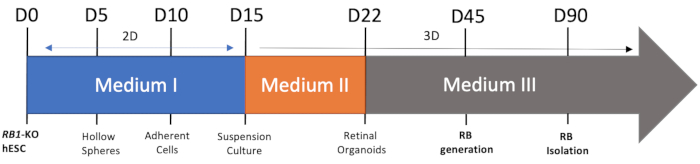
Figure 1: Schematic view of the RB organoids differentiation. Day 0-day 15, the cells are 2D culture in medium I, and after day 15, the cells are suspension culture. RB is formed at around day 45. Please click here to view a larger version of this figure.
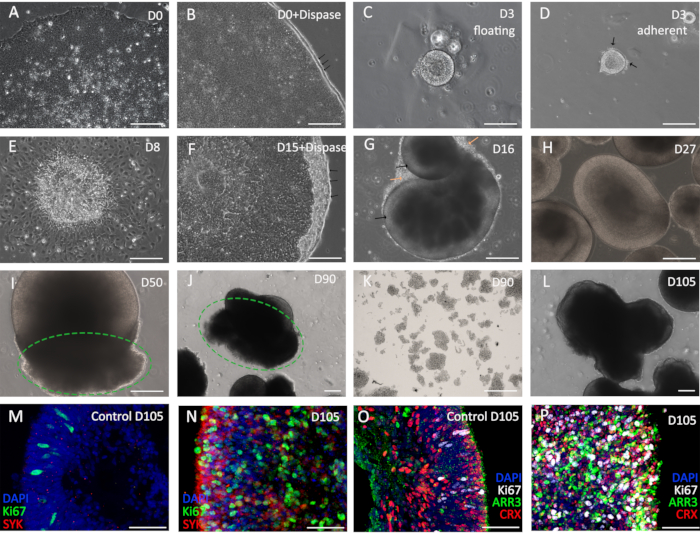
Figure 2: RB generation and characterization. (A–C) The procedure of the early stage, hESC is elevated to form the cysts. The black arrows in (B) show the rolled edges of hESC after dispase treatment. (D,E) The cysts attach to the plates (D) and then expand (E). (F) Adherent cells are elevated to form the retinal organoids. Black arrows indicate the rolled edges after dispase treatment. (G,H) Early days retinal organoids without RB. (I, J) The retinal organoids with RB on day 50 (I) and day 90 (J), the green circles evidence the RB parts. (K) The isolated RB cell line from 90-day retinal organoids. (L) The RB organoids on day 105. (M–P) The immunofluorescence images for the oncogene markers (M,N) and photoreceptor markers (O,P). In A-L, scale bars = 200 µm; in M-P, scale bars = 50 µm. Please click here to view a larger version of this figure.
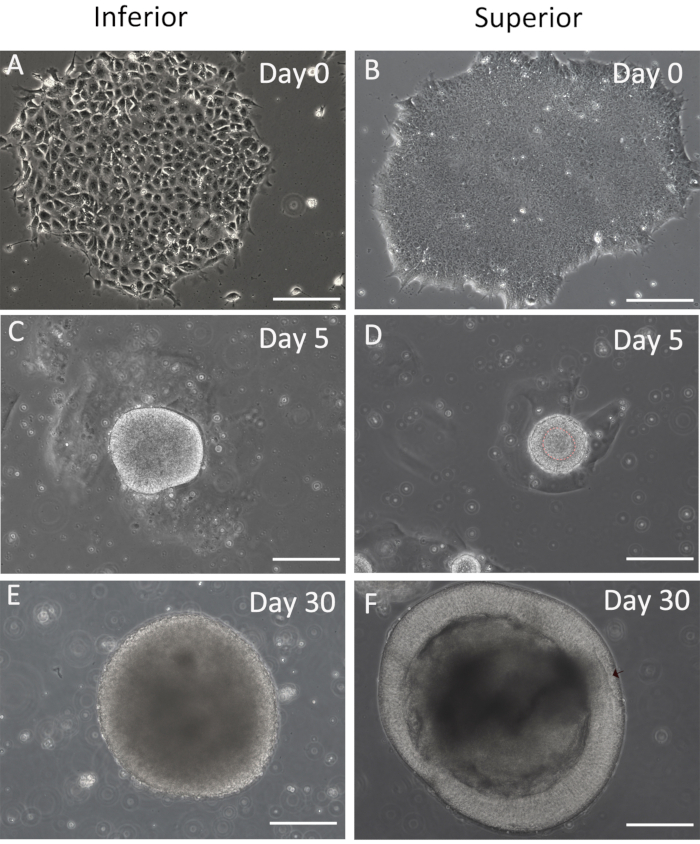
Figure 3: Comparation of the negative and positive results. The inferior and superior images for differentiation on day 0 (A,B), day 5 (C,D), and day 30 (E,F). Scale bars = 200 µm. Please click here to view a larger version of this figure.
